Untitled
The Roles of β-Tubulin Mutations and Isotype Expression
in Acquired Drug Resistance
J. Torin Huzil1, Ke Chen2, Lukasz Kurgan2 and Jack A. Tuszynski11Department of Oncology, University of Alberta, Edmonton, Alberta.
2Department of Computer and Electrical Engineering, University of Alberta, Edmonton, Alberta,
Abstract: The antitumor drug paclitaxel stabilizes microtubules and reduces their dynamicity, promoting mitotic arrest
and eventually apoptosis. Upon assembly of the α
/β-tubulin heterodimer, GTP becomes bound to both the α and β-
tubulin monomers. During microtubule assembly, the GTP bound to β-tubulin is hydrolyzed to GDP, eventually reach-
ing steady-state equilibrium between free tubulin dimers and those polymerized into microtubules. Tubulin-binding
drugs such as paclitaxel interact with β-tubulin, resulting in the disruption of this equilibrium. In spite of several crys-
tal structures of tubulin, there is little biochemical insight into the mechanism by which anti-tubulin drugs target mi-
crotubules and alter their normal behavior. The mechanism of drug action is further complicated, as the description of
altered β-tubulin isotype expression and/or mutations in tubulin genes may lead to drug resistance as has been described
in the literature. Because of the relationship between β-tubulin isotype expression and mutations within β-tubulin, both
leading to resistance, we examined the properties of altered residues within the taxane, colchicine and
Vinca binding
sites. The amount of data now available, allows us to investigate common patterns that lead to microtubule disruption
and may provide a guide to the rational design of novel compounds that can inhibit microtubule dynamics for specifi c
tubulin isotypes or, indeed resistant cell lines. Because of the vast amount of data published to date, we will only pro-
vide a broad overview of the mutational results and how these correlate with differences between tubulin isotypes. We
also note that clinical studies describe a number of predictive factors for the response to anti-tubulin drugs and attempt
to develop an understanding of the features within tubulin that may help explain how they may affect both microtubule
assembly and stability.
Keywords: Tubulin, Microtubule, Isotype, Paclitaxel, Cancer, Resistance, Mutant.
Microtubules (MTs) are large multimeric protein complexes that form hollow cylinders. They are
constructed from repeats of a heterodimer of two 55 kDa proteins known as α and β tubulin. The
formation of the tubulin dimer from the α /β monomers is essentially irreversible, locking a single
molecule of GTP within the non-exchangeable nucleotide binding site of α tubulin (Weisenberg
et al. 1976). Following the formation of the tubulin dimer, an additional molecule of GTP binds the β-subunit and subsequent subunit addition then brings the exposed, nucleotide bound β-tubulin into
contact with α-tubulin from a separate tubulin dimer (Figure 1). This results in the formation of long
chains of tubulin dimers known as protofi laments. This interaction at the α/β tubulin interface, within
the growing protofi laments, promotes the hydrolysis of bound GTP, providing the conformational
fl exibility required during the polymerization-depolymerization cycle (Wang and Nogales, 2005).
In
vitro, these fi laments have been shown to associate laterally, forming sheets which are then thought
to curl in on themselves to form a complete MT. MTs observed
in vitro are composed of 11 to 18
individual protofi laments, whereas most microtubules that are observed
in vivo are made up of only
13 (Unger et al. 1990). MTs are involved in numerous critical, cellular processes including mitosis,
cellular motility, maintenance of cellular morphology, and the activity of cell surface receptors.
(Pichichero and Avers, 1973; Hyams and Stebbings, 1979). Here we discuss the role of MT dynamics
and how tubulin's involvement at the molecular level may affect processes such as chromosome
segregation and cell division. This will be followed by a brief discussion of how this process relates
directly to the chemotherapeutic treatment of cancer and the drugs that are used in this process.
Finally, a discussion of the development of resistance mechanisms to these drugs and how these relate
to tubulin, the expression of isotypes and acquired mutations will hopefully provide insight into the
ever-increasing complexity of this system.
Correspondence: Jack A. Tuszynski. Tel: (780) 432-8906; Email:
[email protected]
Cancer Informatics 2007: 3 159–181
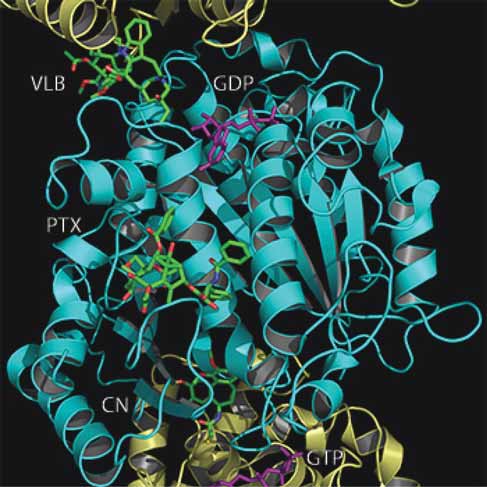
Tubulin structures
like domain to produce crystals, the earliest, 1FFX,
Several crystallographic structures of tubulin as it was derived from the 1TUB data (Gigant et al.
is found within several MT-like conformations are 2000). Two higher resolution structures, 1SA0 and
now available from the RCSB Protein Data Bank 1SA1 followed, producing structures of the colchi-
(PDB) (Nogales et al. 1995; Li et al. 2002; Wang cine and podophyllotoxin bound complexes
and Nogales, 2005). The fi rst tubulin structure, (Ravelli et al. 2004), while both the colchicine and
1TUB, was crystallized as a fl at Zn2+ induced sheet vinblastine binding sites are observed in 1Z2B
of antiparallel protofi lament-like end-to-end α /β (Gigant et al. 2005). Structurally, α and β tubulin
dimer repeats, using docetaxel as a stabilizing are known to be similar, indistinguishable at a
agent (Nogales et al. 1998). Due to diffi culties in resolution of 6 Å, yet share only 40% amino acid
fi tting electron density, this structure contains identity (Li et al. 2002). Each tubulin monomer
misalignments and was superseded by 1JFF, in can be divided into three distinct domains. The
which paclitaxel was utilized as a stabilizing agent amino terminal domain (composed of residues
(Lowe et al. 2001). Similarly, a third structure 1–205), an intermediate domain (residues 206–
1TVK uses epothilone A, which binds at the same 381) and a carboxy terminal domain, which
taxane binding site, stabilizing the MTs (Nettles contains the flexible C-terminal tail (residues
et al. 2004). Additional structures use a stathmin-
382–444). Unfortunately, while there is a wealth
Palitaxel, Colcicine and Vinblastine Binding Sites
Figure 1. Paclitaxel, colchicine and Vinca binding sites on α /β tubulin protofi lament. Shown here is a cartoon representation of a protofi la-
ment with superimposed drug molecules, shown as space fi lling spheres. Structures of the paclitaxel (PTX), colchicine (CN) and vinblastine (VLB) from structural fi les 1JFF, 1SA0 and 1Z2B have been superimposed and fi t back onto the 1SA0 structure to obtain the relative posi-tioning of each drug within the protofi lament. A single α /β-tubulin heterodimer comprises the β tubulin monomer (cyan) in the center of the
frame and two α tubulin monomers (yellow) at the top and bottom of the frame. The GTP at the non-exchangeable and GDP at the exchange-
able site are colored purple.
Cancer Informatics 2007: 3
The Roles of β-Tubulin Mutations and Isotype Expression in Acquired Drug Resistance
of information on the structure of tubulin, little is (Margolis and Wilson, 1981; Horio and Hotani,
known about the functional signifi cance related to 1986; Melki et al. 1989; Walker et al. 1991;
its structural features. This is almost entirely due Mandelkow and Mandelkow, 1992; Zheng et al.
to the lack of molecular understanding of the 1995). MTs comprise much of the mitotic spindle
dynamic properties of tubulin dimers and how they apparatus and its proper assembly is essential, as
behave within microtubules.
it likely provides the substantial mechanical force
for mitotic chromosome segregation (Westermann
et al. 2006). Assembly failure of the mitotic spindle
Microtubule structure and dynamics
generally results in mitotic arrest, apoptosis, and
and the effect of drug interactions
eventually cell death. As such, MTs have become
MTs exhibit an irregular temporal pattern of the target for a large number of anti-mitotic agents
assembly and disassembly, which has been termed including antitumor drugs such as the taxanes,
dynamic instability (Mitchison and Kirschner, epothilones, colchicine and Vinca alkaloids,
1984). The process of dynamic instability features including vinblastine and vincristine (Jordan and
episodes of distinct catastrophes that result in MT Wilson, 2004).
collapse, and rescues that lead to microtubule re-
The method of action of these drugs is to either
growth (Bayley et al. 1990; Drechsel et al. 1992; promote or inhibit MT polymerization by binding
Odde et al. 1995). The assembly and disassembly at specific sites on the surface of α/β-tubulin
processes of MTs, both in vivo and in vitro, have heterodimers (Figure 1). Paclitaxel, which
been extensively studied and are regulated by continues to be one of the most successful cancer
mechanisms that are sensitive to temperature, pH therapeutic agents, has a unique mechanism of
and ionic concentrations that can accommodate the action as it binds to and results in the stabilization
critical roles played by MTs at different phases of of MTs within all cells (Wani et al. 1971; Schiff
the cell cycle. Following the initial nucleation of and Horwitz, 1980). Derivatives of paclitaxel, such
a MT, the addition of subsequent tubulin dimers as docetaxel, have been synthesized to address the
can occur at either of its free ends. Both polymer-
limited solubility of paclitaxel and show increased
ized and unpolymerized tubulin monomers can binding to β-tubulin (Ringel and Horwitz, 1991).
bind free GTP, and upon assembly, the energy of Two additional families of similar tubulin-binding
hydrolysis from GTP to GDP is imparted into the drugs are the epothilones and dolastatins, which
tubulin subunits, however the outcome of this share a similar mode of binding. Interest in the
process and the fate of the energy released is yet epothilones over existing MT stabilizing agents
unknown. It has been hypothesized that when β- such as paclitaxel is primarily due to their ability
tubulin becomes bound within a MT, the exchange to retain cytotoxicity in multidrug resistant (MDR)
of bound GDP for GTP is blocked, and as a result, cells (Bollag et al. 1995). Compounds, such as
the protofi lament maintains its structure as long as discodermolide and sarcodictyin have been shown
there is not an exposed β-tubulin at the growing to inhibit the proliferation of human cells through
end. Stabilization of the entire MT structure is a mechanism similar do that of the taxanes. These
normally afforded by the binding of a GTP cap at compounds seem to selectively stabilize MTs
the end of the MT which shields the terminal β during cell division, and outcompete paclitaxel for
tubulin from a conformational change that is then tubulin binding (Klein et al. 2005). In the absence
thought to induce its dissociation (Mitchison and of paclitaxel, it has been predicted that the energy
Kirschner, 1984). We believe that at least part of stored in longitudinal contacts is greater than that
the relatively large amount of energy is stored stored in lateral contacts (VanBuren et al. 2002).
within the MT itself in the form of potential confor-
Recent results using hydrogen/deuterium exchange
mational energy. When a critical amount of this coupled with mass liquid chromatography
potential energy has been exceeded, it is released have demonstrated that increased rigidity in
in the form of a catastrophic collapse of the entire paclitaxel stabilized MTs was distinct from stabi-
lization as a result of GTP-induced polymerization
The dynamic properties of MTs are especially (Xiao et al. 2006). The authors suggest that pacli-
critical during mitosis, when a delicate equilibrium taxel increases the overall energy associated with
of distances and forces is necessary for proper polymerization while maintaining the same differ-
chromosome alignment prior to segregation ential between lateral and longitudinal contacts in
Cancer Informatics 2007: 3
the microtubule lattice. Through structural studies develop a drug or treatment regimen that will target
both the taxanes and epothilones were shown to only cancer cells and will target them selectively.
bind a unique site within the β subunit of the α /β- In the case of paclitaxel, this would require deter-
tubulin heterodimer (Lowe et al. 2001; Nettles et mining the differences between tubulin as observed
al. 2004). Unfortunately, neither of the structures in cancerous cells and non-cancerous cells. Fortu-
has been able to reveal the precise mechanism of nately there are slight differences within several
MT stabilization by these drugs.
β-tubulin isotypes expressed in a range of cell types
The Vinca alkaloids include vincristine vinblas-
that may afford a foundation for the development
tine and vinorelbine and are used most commonly of anti-tubulin drug derivatives with increased
in combination chemotherapy regimes. Vinblas-
specifi city for particular cancer cells. Unfortu-
tine was shown to bind at the inter-tubulin dimer nately, as the precise binding modes of these
interface, ultimately resulting in the net reduction compounds within β-tubulin is not yet able to fully
of polymerized tubulin concentration (Gigant explain how these drugs affect MT stabilization,
et al. 2005). At high concentrations the Vinca our ability to create a precise molecular model of
alkaloid, vinblastine, binds to MTs and results in their effects is limited.
their depolymerization. However, at low concen-
trations, vinblastine is thought to bind to MT tips
and suppresses their dynamic instability, leading Tubulin isotypes
to stabilization (Toso et al. 1993). We feel that this Many eukaryotic organisms carry multiple
may be a result of vinblastine's ability to maintain genomic copies of functional α or β tubulin,
the tubulin dimer at the end of the growing MT in commonly referred to as isoforms (or isotypes if
a slightly bent conformation (Wang and Nogales, they are confi ned to a single organism). At the
2005). If the tip of a protofi lament was maintained cellular level, the role of tubulin is extremely
with a slightly bent conformation, due to the posi-
complex and seems to be related to subtle struc-
tioning of vinblastine between the dimers, then at tural variations observed between the α and β
low enough concentrations, a small amount of isoforms (Richards et al. 2000). For instance, MT
drug bound to the end of the MT would mimic the dynamics appears to change signifi cantly as a
presence of a GTP cap, thereby resulting in MT function of β tubulin isotype expression (Panda
et al. 1994). This result implies that if MT assembly/
The third observed drug site within β-tubulin disassembly equilibrium is disrupted through some
was shown to bind colchicine, a water-soluble external stimulus, cells could respond by producing
alkaloid that, like paclitaxel, binds to α /β tubulin an appropriate isotype mix to restore normal
dimers and blocks cell division thereby inhibiting balance. The existence and varied distribution of
mitosis (Ravelli et al. 2004). Unlike paclitaxel or tubulin isoforms provides a link to their function in
vinblastine at low concentration, the binding of the polymerization and stability of MTs. While the
colchicine does not result in MT stabilization, but presence of numerous tubulin isoforms suggests that
instead results in their destabilization since colchi-
they may play specifi c roles in MT function, there
cine binds only to free tubulin and not to tubulin are no precise predictive models to describe differ-
polymerized into MTs.
ences between them.
Following exposure to all of these drugs, cells
In humans, several α and β tubulin isotypes have
experience mitotic arrest, which eventually leads been identifi ed and characterized (Lu and Luduena,
to apoptosis. This makes these drugs extremely 1994; Luduena, 1998; Roach et al. 1998). While α
effective chemotherapeutic agents' for targeting tubulin must play an obvious role in the determination
all rapidly dividing cancerous cells. Nevertheless, of MT function, we have chosen to focus only on β
even the most successful chemotherapy drugs have tubulin for this discussion, as most of the available
undesirable side effects that limit their utility. The data in the literature deals with this protein as a target
anti-tubulin chemotherapeutic agents' main fl aw for drug action and protein-protein interactions.
is that they bind tubulin indiscriminately, leading Through a search of available protein sequence data-
to the widespread destruction of both cancerous bases, we have previously identifi ed a total of ten
and healthy cells, resulting in unwanted side-
unique β tubulin isotypes, all of which have related
effects such as neuropathy (Rowinsky et al. 1989). amino acid sequences and are generally well
The ultimate goal of chemotherapy research is to conserved (Huzil et al. 2006). Despite having similar
Cancer Informatics 2007: 3
The Roles of β-Tubulin Mutations and Isotype Expression in Acquired Drug Resistance
Figure 2. Sequence alignment of β tubulin isoforms. Each of the ten Human β-tubulin isoforms that was identifi ed in our screen of the Uni-
prot and NCBI Entrez databases was aligned using the ClustalW software package (Thompson et al. 1994). Prior to performing the alignment, the highly variable carboxy terminal residues were removed from each sequence. This was done as the structural fi le, 1JFF, does not con-tain any of these residues. At each position within the alignment, black boxes indicate identical residues, grey boxes indicate residues that are conserved, while white boxes indicate residues that are divergent. The 6 Å cutoff binding sites for paclitaxel, colchicine and vinblastine are indicated by boxes labeled PTX, CN and VLB, respectively.
Cancer Informatics 2007: 3
sequences, specifi c regions of higher sequence vari-
in cellular components that interact with the target.
ability have been identified (Figure 2). The highest Cell culture studies have previously demonstrated
degree of sequence variability between the α and that the most frequent mechanism for resistance to
β-tubulin isotypes occurs in the extreme carboxy drugs such as the colcemides and Vinca alkaloids
terminal region (residue 430 and greater). This is P-glycoprotein mediated multidrug resistance
C-terminal region has been used to identify distinct (Cabral and Barlow, 1989; Dumontet et al. 1996).
β-tubulin isotypes, based on their reactivity with However, in direct contrast to this observation,
monoclonal antibodies (Banerjee et al. 1990; Panda cells which show resistance to taxanes tend to
et al. 1994).
exhibit alterations in tubulin expression patterns
Each β-tubulin isotype seemingly has a unique with low P-glycoprotein activity. In addition to the
pattern of expression ranging from highly specifi c altered expression of β-tubulin isotypes, point
for classes III, IVa and VI, to constitutive expres-
mutations in tubulin leading to alterations and
sion for classes I and IV (Cowan and Dudley, 1983; expression related post-translational modifi cations
Jensen-Smith et al. 2003). Class I β-tubulin is the of tubulin regulatory proteins, such as stathmin,
most commonly expressed isotype in humans and microtubule associated protein (MAP), tau and
as such is also the most common isotype found in MAP4 have also been implicated in changes to MT
cancer cells (Sullivan and Cleveland, 1986). Both dynamics and the development of drug resistance
classes II and III β-tubulin have been observed at (Liu et al. 2001).
increased levels in human tumors (Scott et al. 1990;
In addition to isotype expression and acquired
Ranganathan et al. 1996; Kavallaris et al. 1997; mutations, tubulin is also thought to undergo an
Prasannan et al. 2000; Dozier et al. 2003; Ferguson auto-regulatory mechanism where treatment with
et al. 2005; Mozzetti et al. 2005).
MT destabilizing drugs, such as colchicine, will
Recently, the role of β-tubulin isotypes in resis- ultimately result in the coordinated degradation of
tance to anti-mitotic drugs has become a topic of tubulin mRNA and a corresponding reduction of
great interest, as the design of novel drugs based cellular free tubulin dimers. The synthesis of α and
on isotype specifi c differences would result in β-tubulin monomers appears to be regulated by a
better treatment protocols (Burkhart et al. 2001). mechanism in which α-tubulin translation is inhib-
However, in addition to functional tubulin ited by free α-tubulin but not by α-tubulin that is
isotypes, the tubulin gene family also contains complexed with the β-subunit (Gonzalez-Garay
several pseudo-genes, which are non-functional and Cabral, 1996). In this way, β-tubulin is synthe-
sequences within the genome having close simi-
sized only when there are available α-subunits to
larities to functional genes. There are several which it can bind. The treatment of cells with MT
human β-tubulin pseudo-genes, all of which show stabilizing drugs, such as paclitaxel results in the
substantial homology to the Class I β-tubulin gene, overall stabilization of tubulin mRNA and an
a situation which has resulted in the erroneous increase in tubulin protein due to a net reduction
identifi cation of several tubulin mutations that in concentration directly following drug treatment.
correlate with paclitaxel resistance (reviewed by This results in the overall equilibrium of free
Berrieman et al. 2004). This has added controversy tubulin within the cell and maintains a steady
to an otherwise attractive hypothesis that muta-
supply of tubulin for MT synthesis (Boggs and
tions within the β-tubulin genes can lead to drug Cabral, 1987; Theodorakis and Cleveland, 1992;
Barlow et al. 2002). This further implies that any
mutation or expression of a tubulin isotype that
affects the overall stability of MTs will also have
Drug resistance mechanisms
an effect on the total amount of free tubulin dimers
involving tubulin
that are present within the cell.
As with all chemotherapeutic treatments, the devel-
Finally, tubulin can also be controlled post-
opment of drug resistance to anti-tubulin agents is translationally through degradation. Although
a major clinical problem with no simple solution. tubulin heterodimers are very stable (Spiegelman
Drug resistance can originate through several et al. 1977), excess free subunits (Gonzalez-Garay
mechanisms involving: (a) changes in cellular drug and Cabral, 1995) or defective tubulin proteins
uptake, (b) drug metabolism, (c) structural changes have a much shorter half-life (Kemphues et al.
in the drug target, (e) drug effl ux, or (f ) changes 1982; Boggs and Cabral, 1987). The tubulin
Cancer Informatics 2007: 3
The Roles of β-Tubulin Mutations and Isotype Expression in Acquired Drug Resistance
molecule is also subject to several post-transla-
isotypes demonstrates that many of the differences
tional modifi cations which are thought to also between them occur outside the main drug binding
affect their rate of assembly into MTs. Some of sites (Figure 2). When displayed on the surface of
these modifi cations, such as phosphorylation and the 3D tubulin structure, it becomes apparent that
acetylation are common to many proteins. Others, most of the differences found between the various
such as polyglutamylation, are very rare, while still isotypes are restricted to the lateral and longitudinal
others, such as tyrosinolation/detyrosinolation and surfaces that are involved in protein-protein inter-
polyglycylation, have so far been reported only in actions with the neighboring tubulin dimers in the
tubulin (Luduena, 1998; MacRae, 1997; Wester-
MT (Figure 3). Differences at these locations may
mann and Weber, 2003; Soucek et al. 2006).
provide the cell with the ability to alter expression
patterns for tubulin isotypes and in this way affect
the overall kinetics of MT assembly and disas-
Drug resistance due to β-tubulin
sembly. This could be in response to such external
isotype expression levels
stimuli as temperature changes or drug exposure.
Several studies have pointed to the over-expression Interestingly, several differences can be observed
of tubulin isotypes as a possible mechanism on the exterior face of the β-tubulin when it is
resulting in drug resistance ( Banerjee et al. 1990; assembled into the MT, while there are signifi cantly
Minotti et al. 1991; Banerjee and Luduena, 1992; fewer observed differences on the inner surface.
Haber et al. 1995; Kavallaris et al. 1997; Derry This may be a result of selective differences
et al. 1997; Banerjee and Kasmala, 1998; Ranga-
between isotypes that are meant to accommodate
nathan et al. 1998; Verdier-Pinard et al. 2003). Microtubule Associated Protein (MAP) binding,
While other studies suggest that expression of thereby leading to altered MT kinetics (Kurz and
certain tubulin isotypes does not lead to paclitaxel Williams, 1995).
resistance (Blade et al. 1999; Nicoletti et al. 2001;
The underlying molecular mechanisms of
Sale et al. 2002). However, as more data is tubulin isotype expression, in particular Class III
acquired, it is becoming clear that isotype expres-
β-tubulin over-expression, in drug resistance are
sion does lead to drug resistance at some level. In not well understood. There are a number of possi-
particular, Class III β-tubulin has been implicated bilities, including tubulin's normal vibrational
in paclitaxel resistance through a mechanism that modes, perhaps resulting in altered protein
results in decreased MT stability, thus counter-
dynamics. More likely, it is a critical change
acting the effect of paclitaxel (Sangrajrang et al. within surfaces that interact during MT assembly.
1998; Katsetos et al. 2003; Hari et al. 2003a; Shalli Therefore, the expression of specific tubulin
et al. 2005). More specifi cally it appears that the isotypes could result in the alteration of MT
expression of specifi c tubulin isotypes, such as stability. A cellular requirement for less stable
βIII, may not affect MT dynamic instability in the MTs may then result in the over-expression of
absence of drugs such as paclitaxel, but may act to isotypes such as βIII, which has been demon-
suppress the drug's ability to affect MT dynamics strated to be produce more dynamic MTs than
directly (Kamath et al. 2005). Finally, support for other isotypes (Derry et al. 1997).
the role of tubulin isotype expression and their
effect on drug resistance was observed when anti-
sense oligonuclotides specifi c for βIII result in the Tubulin mutations
resensitization of resistant cells to paclitaxel and drug resistance
(Kavallaris et al. 1999). In addition to the several The fi rst studies that identifi ed resistance to anti-
observations of tubulin isotype expression patterns mitotic drugs through a mechanism of β tubulin
in tumor cells, the expression of tubulin isotypes mutation were originally performed in the early
has also been implicated in the differentiation state 1980's (Ling et al. 1979; Cabral et al. 1980;
of cells (Carles et al. 1999).
Keates et al. 1981; Cabral et al. 1982; Cabral and
We have previously described the construction Barlow, 1989). However, the work of Monzo
of a complete set of homology models for the et al. (1999) in observing mutations of β-tubulin
human β-tubulin isotypes and described several in human lung cancer cell lines has been the
differences that we observed between them (Huzil source of much controversy since its publication.
et al. 2006). Sequence alignment of all the β-tubulin It has since been suggested that the presence of
Cancer Informatics 2007: 3
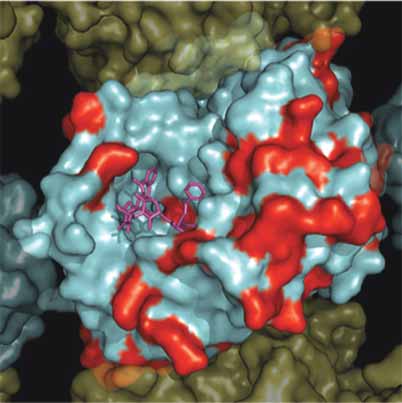
A. β-Tubuiln Isotypes MT Interior
B. β-Tubuiln Isotypes MT Exterior
Figure 3. Surface map of nonconserved residues in all Human β-tubulin isoforms. A solvent accessible surface was drawn onto the α /β-
tubulin heterodimer obtained by Nogales et al. 2001 (PDB identifi er 1JFF). This fi gure shows an α /β-tubulin heterodimer within a MT contain-
ing 13 protofi laments which was reconstructed as described by Li et al. (2002). The α-tubulin surface is colored yellow and the β-tubulin
surface colored cyan with nonconserved positions colored red. Panel A. Illustrates the MT from the interior surface and shows the binding
site for paclitaxel (purple). Panel B. Illustrates a 180° rotation about the y-axis to show the exterior surface of the microtubule.
Cancer Informatics 2007: 3
The Roles of β-Tubulin Mutations and Isotype Expression in Acquired Drug Resistance
tubulin pseudo-genes and sequencing errors result of reported mutations that confer resistance to
in erroneous interpretation and therefore makes tubulin-binding drugs in all reported mammalian
determining true mutations in tubulin diffi cult. cells (Table 1). There seemingly is little correlation
Many sequencing errors within the β-tubulin gene between the overall structure of the paclitaxel
tend to be a result of primer selection where binding site and the location of mutations in β-
intronic primers are used or pseudo-genes are tubulin that occur as a result of resistance following
inadvertently sequenced (de Hasegawa et al. exposure to paclitaxel (Figure 4). We have observed
2002; Tsurutani et al. 2002; de Castro et al. 2003; that within the set of 21 unique positions within
Achiwa et al. 2003; Maeno et al. 2003; Berrieman the tubulin sequence, only eleven mutations were
et al. 2004; Mesquita et al. 2005).
found to be contained within the taxane binding
It has been suggested that differences in drug site (Figure 5a). Only eight of these substitutions
binding affi nities do not play a signifi cant role for were identifi ed as becoming either paclitaxel or
the colcimides or vinblastine with regards to the epothilone resistant. Interestingly three of the
acquisition of tubulin mutations in certain cell lines mutations within the taxane binding site resulted
(Hari et al. 2003b). However, there are several in MT stabilization and as a result became resistant
studies, discussed below, that point to a role for to either colchicine or vinblastine. Of the remaining
mutations within the taxane binding site as a ten sites within β-tubulin, fi ve were shown to
mechanism for resistance to paclitaxel. We feel that become paclitaxel resistant, yet did not occur in
mutations observed within the β-tubulin genes may the vicinity of the taxane binding site (Figure 5b).
not result in changes to drug binding affi nity, but Of two sites found near the colchicine binding site,
will more likely play a role at distant locations only K350N was identifi ed as being colchicine
within the protein. The rarity of true somatic muta-
resistant (Figure 6a) (Hari et al. 2003b). Interest-
tions in tubulin in clinical samples also suggests ingly, mutation L240I, which is also found near
that other mechanisms, such as drug effl ux or the colchicine binding site, confers resistance to
changes in the expression levels of the different vinblastine (Kavallaris et al. 2001). This was a
β-tubulin isotypes, are a more important contrib- remarkable observation as the Vinca binding site
uting factor for anti-tubulin drug resistance (de is on the opposite face of β-tubulin from the colchi-
Castro et al. 2003; Maeno et al. 2003; Hasegawa cine binding site (Figure 1). There are also two
et al. 2002; Ferguson et al. 2005). However, the mutations situated near the Vinca binding site. The
positive data obtained with cell lines suggests that first at S172 conferred vinblastine resistance
further investigation is needed to ascertain what (Poruchynsky et al. 2004) and the second, at P173
role, if any, mutations in the β-tubulin gene have curiously conferred epothilone resistance with an
in predicting clinical response to anti-tubulin opposing MT destabilizing phenotype (Figure 6b)
agents. We cannot discount the role of intronic (He et al. 2001).
mutations and their observed role for drug resis-
tance and how this may ultimately affect gene Mutations found within the taxane
expression of a particular tubulin isotype as de
Castro et al. (2003) observed that as alterations in binding site
β-tubulin were amplifi ed with exonic primers less When examining mutations acquired following
clinical response to paclitaxel was observed. Addi-
exposure to taxanes or epothilones, several are
tionally, observations made with regards to found directly within the taxane binding site and
acquired mutations are often complicated by the could therefore affect drug binding through VDW
concurrent overexpression of different tubulin (Van der Waals (VDW) interactions, electrostatics
isotypes, making the interpretation of results or hydrophobic differences. These include C211,
L215, L217, D224, L228, A231, S234, F270, T274
and R282 A364 (Figure 5a). We should note that
while all of these residues are located within close
Analysis of Specifi c β-Tubulin
proximity of the bound paclitaxel molecule from
the 1JFF structure, most were reported not to
To determine which residues within each of previ-
change the affi nity of the taxanes or epothilones,
ously described drug binding sites within β-tubulin but appear to destabilize MTs in the absence of any
are essential for activity, we examined the location drug (Gonzalez-Garay et al. 1999; Verrills et al.
Cancer Informatics 2007: 3
Table 1. Observed β tubulin mutations in mammalian cell lines as a result of acquired drug resistance. The
columns representing drug interactions indicate residues in which any atom is within 6 Å of any atom within the
bound drug molecule from structural fi les 1JFF, 1SA0 and 1Z2B for Paclitaxel (PTX), Colchicine (CN) and Vin-
blastine (VLB) respectively.
Drug Interactions
Mutation Cell
Type Reference
Phenotype
Barlow 2002, Hari
Increased MT stabilization
Growth dependent on Colchicine
Wang 2004, Yang 2005 Decreased MT stabilization
Taxol resistant Cis-acting suppressor
Poruchynski 2004
Increased MT stabilization
Hemiasterlin Colchicine and
Vinca resistance.
Decreased MT stabilization
Epothilone A resistant.
Increased MT stabilization
2-Methoxyestradiol and Vinca
resistant. No effect on Taxol
or Colchicine phenotype
Increased MT stability
Colchicine and vinca
Gonzales 1999, Barlow Decreased MT stabilization
Taxol resistance.
Not implicated in drug binding
Normal Phenotype
Decreased MT stabilization
Taxol sensitivity, but not
Epothilone Sensitive
Decreased MT stabilization
Taxol resistant. Not implicated
Increased MT stability
Colchicine and vinca resistance
Decreased MT stabilization
Taxol resistance. Not
implicated in drug binding
Decreased MT stabilization
Taxol resistant, hypersensitive
to destabilizing compounds
Increased MT stability
Colchicine and vinca resistance
Increased MT stabilization
Vinca and Colchicine resistant
Cancer Informatics 2007: 3
The Roles of β-Tubulin Mutations and Isotype Expression in Acquired Drug Resistance
Drug Interactions
Mutation Cell
Type Reference
Phenotype
Giannakakou 1997
Decreased MT stabilization
Taxol resistance
Giannakakou 2000
Decreased MT stabilization
Epothilone resistant reduced
sensitivity to Taxol may
affect drug binding
Giannakakou 2000
Decreased MT stabilization
Epothilone resistant reduced
sensitivity to Taxol may
affect drug binding
He 2001, Verrills 2003, Decreased MT stabilization
Wang 2004, Yang 2005 Taxol resistant Cis-acting
suppressor of D45Y
Not known how this confers
only a genetic study
Hari 2003,Gokmen
Increased MT stabilization
2-Methoxyestradiol and vinca
resistant. No effect on Taxol
or Colchicine phenotype
Giannakakou 1997
Decreased MT stabilization
Decreased MT stabilization
Epothilone A resistant
2003; Hari et al. 2003b). Of these eleven residues that both the paclitaxel and colchicine binding sites
found to be directly in contact with either paclitaxel are nearby, but also because many mutations
or epothilone, only F270V, T274I and R282N were conferring resistance to drugs that either promote
reported to have a direct effect on drug binding or inhibit MT assembly are also located in this
affi nity (Giannakakou et al. 1997; Giannakakou et same area (see following discussion). This is an
al. 2000). This observation suggests that there must interesting result, as these residues are clearly
be a global effect on tubulin dimer interactions that located near the taxane binding site and are within
are involved in MT assembly rather than a reduced 8–10 Å of bound paclitaxel. However, their prox-
paclitaxel binding effect as the mechanism for drug imity to both the GTP binding site and the colchi-
cine binding site may also result in alternate
C211/D224/S234. Substitutions at C211, D224 phenotypes depending on the substitutions. This
and S234 all were all shown to result in resistance is supported by the observation that mutations at
to both colchicine and vinblastine, producing an these positions in yeast were shown to confer vari-
increase in MT stability and the amount of polym-
able drug resistance depending on the amino acid
erized MT observed within cells (Hari et al. 2003b). substitution indicating the critical role that this
All of these substitutions are located within a region plays in MT stability and drug resistance
structure known as the M loop, which has been (Gupta et al. 2001).
shown to be critical for the function of paclitaxel
L215, L217, L228. Analysis of β-tubulin alleles
and MT dynamics, affecting lateral interactions from nine paclitaxel-resistant Chinese Hamster
between tubulin subunits in the MT (Figure 5a) Ovary (CHO) cell lines revealed a cluster of muta-
(Keskin et al. 2002). The signifi cance of this region tions affecting L215, L217, and L228 (Gonzalez-
in MT assembly is illustrated, not only by the fact Garay et al. 1999). At position 215, histidine,
Cancer Informatics 2007: 3
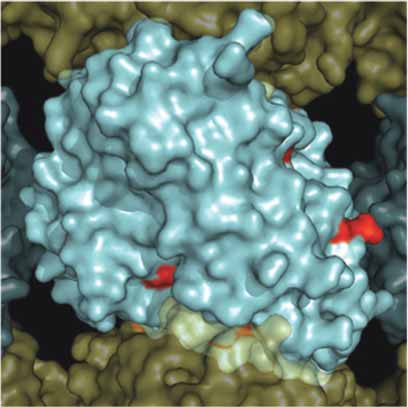
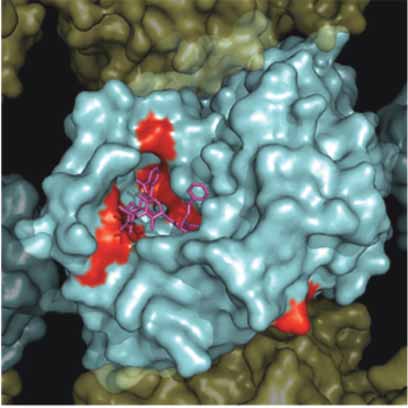
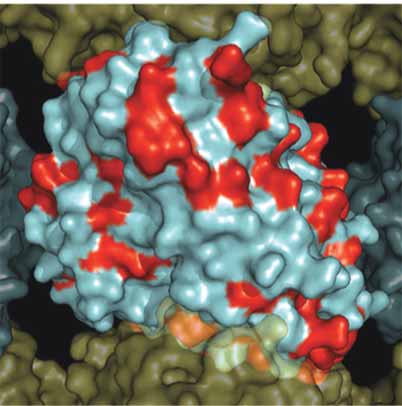
A. β-Tubuiln Mutations MT Exterior
B. β-Tubuiln Mutations MT Exterior
Figure 4. Surface map of mutation sites in mammalian β-tubulin. As in Figure 3, a solvent accessible surface was drawn onto the α /β-tubu-
lin heterodimer and assembled into a 13 protofi lament MT. Here the β-tubulin surface is colored red to illustrate the surface positions of
mutations observed in Table 1. Panel A. Illustrates the MT from the interior surface and shows the binding site for paclitaxel (purple).
Panel B. Illustrates a 180° rotation about the y-axis to show the exterior surface of the microtubule.
Cancer Informatics 2007: 3
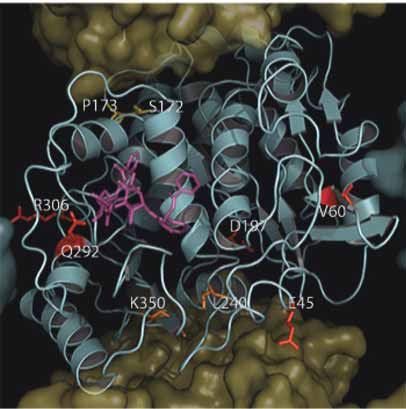
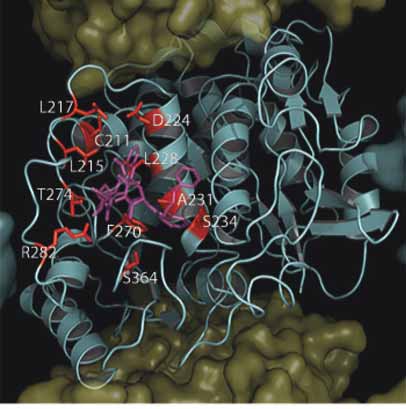
The Roles of β-Tubulin Mutations and Isotype Expression in Acquired Drug Resistance
A. β-Tubuiln Mutations Within PTX Site
B. β-Tubuiln Mutations Not In PTX Site
Figure 5. Location of mutations in mammalian β-tubulin in the taxane binding site. As in Figure 3, with the exception that the β-Tubulin surface
has been removed to expose the underlying tertiary structure which is shown as a cyan cartoon. Panel A. Shows mutations found within the
taxane Binding Site (C211, L215, L217, D224, L228, A231, S234, T274, F270, R282, S364) colored red in relation to paclitaxel (purple).
Panel B. Shows residues found outside the taxane binding site (E45, V60, Q292, R306) colored red. Residues found within the Vinca binding
site (S172 and P173) are colored yellow and residues found within the colchicine binding site (L240 and K350) are colored orange.
Cancer Informatics 2007: 3
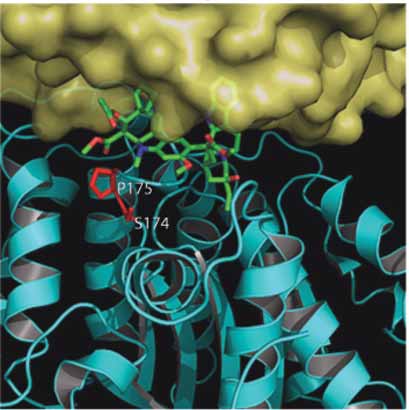
A. Colchicine Binding Site Longitudinal Surface
B. Vinblastine Binding Site Lateral Surface
Figure 6. Location of mutations in mammalian β-tubulin in the colchicine and Vinca binding sites. As in fi gure 5, the surface has been removed
from β-tubulin. Panel A. Shows a view of the colchicine binding site from the longitudinal surface between an α /β-tubulin heterodimer. The
α-tubulin has been removed to facilitate viewing of the bound colchicine within the β-tubulin a second α-tubulin can be seen behind
β-tubulin in the background. Colchicine is colored green and is shown as sticks. The two mutated residues (L240 and K350) are shown as
red sticks. Panel B. Shows a view of the Vinca binding site from the lateral surface between two protofi laments. The vinca alkaloid vinblas-
tine is shown as green sticks and the two mutated residues (S174 and P175) are shown as red sticks.
Cancer Informatics 2007: 3
The Roles of β-Tubulin Mutations and Isotype Expression in Acquired Drug Resistance
arginine, or phenylalanine substitutions were
The C4–C5 oxetane of the bound taxane is
observed, while at position L217, an arginine and located near the polar backbone atoms of residues
at L228 either histidine or phenylalanine substitu-
273, 275, 276 and the hydroxyl side chain of T274.
tions were observed. All of these mutations resulted Therefore, substitutions of this residue may lead
in a decrease in MT stability, as demonstrated by to a change in the hydrophillicity of this region
an overall reduction in the level of polymerized making taxane binding less favorable. Addition-
tubulin within the cell and by increased resistance ally, Giannakakou et al. (2000) also hypothesized
to paclitaxel. Substitutions at positions L215 and that cross-resistance to both epothilone and pacli-
L217 are in a loop that connects helices H6 and taxel may be a result of the epothilone C7-OH
H7, while L228 is located within H7 itself hydrogen bond with the vicinity of T274. There-
(Figure 5a). These substitutions may produce fore, T274I results in the loss of this hydrogen
resistance as a result of their location in or near H7 bond. In contrast, paclitaxel has hydrogen bond
since it has been hypothesized that H7 controls the donors or acceptors at their C10, C9, and C7
conformation of the entire molecule (Amos and positions and can presumably form alternate
Lowe, 1999). An interesting observation was made hydrogen bonding patterns.
by Wang et al. (2006) when the substitution L215V
F270V, A364T. Cell cultures resistant to pacli-
did not produce any observable phenotype while taxel but not epothilone and not dependent on
mutation to any other residue produced cells resis-
paclitaxel for growth, suggested that the mutation
tant to paclitaxel as a result of global MT destabi-
F270V is specifi c only for paclitaxel binding and
lization. Perhaps the specifi c spatial arrangement does not lead to an overall reduction in MT
of the two methyl groups found on leucine and assembly kinetics (Giannakakou et al. 2000).
valine results in no observable effect for this muta-
However, collateral sensitivity to vinblastine
tion. However, this observation requires additional suggests a global destabilization of the MT struc-
experimentation before a concrete mechanism can ture also. This mutation was independently selected
be established for this apparent selectivity.
along with another mutation, A364T, which also
T274/R282. In addition to describing a common demonstrated the same paclitaxel, but not epothi-
pharmacophore for Epo A/Epo B binding which lone-resistant phenotype. Both of these substitu-
may explain mutational effects at L215, L217 and tions occur within the taxane binding site and
L228, Giannakakou et al. (2000) also identifi ed comprise the majority of the surface that makes up
mutations at positions T274 and R282 as conferring the fl oor of this binding site (Figure 5a). Several
either epothilone or paclitaxel resistance. Both of possible interactions with the taxane C13 side chain
these mutations resulted in a similar reduction in are possible. Substitution of A364T would result
the overall amount of polymerized tubulin within in a polar residue in this normally hydrophobic
the cell. While these mutations did seem to have an surface. While F270 does not seem to participate
effect on the ability of tubulin to bind either epothi-
in any signifi cant ring interactions, this effect
lone or paclitaxel, they also conferred a greater cannot be ruled out as it is in close proximity to
degree of MT destabilization, which is most likely both C13 phenyl rings. The resistance to paclitaxel
the major factor leading to drug resistance. Both of but not epothilone for both of these substitutions
these residues are found within the M loop and their suggests that there is a difference in the binding
interactions with the taxane site are direct and most pocket that does not affect epothilone binding. This
likely signifi cant (Figure 5a). As the amino terminus effect may be due to the epothilone molecule sitting
of the M loop comprises part of the taxane higher within the binding pocket at this region
ring-binding region, a mutation of R282 could and/or due to the lack of potential aromatic interac-
directly affect the binding of both the taxanes and tions with F270.
epothilones. However, as previously discussed,
A231. Through screening a series of desoxye-
the involvement of the M loop within longitudinal pothilone B resistant leukemia cell lines, the muta-
MT interactions is extremely important for proper tion A231T was seen to be producing less stable
assembly and stability. The importance of MTs (Verrills et al. 2003). This substitution was
this region is further supported by the observation shown to confer resistance to the epothilone analog
that both T274 and R282 are evolutionarily desoxyepothilone B, but did not seem to alter drug
conserved in all known human β-tubulin isotypes binding. The authors also observed that cells
carrying this mutation were hypersensitive to
Cancer Informatics 2007: 3
microtubule destabilizing agents and expressed lizing drugs, such as paclitaxel or epothilone, to
increased levels of class III β-tubulin. These cells compensate for the increased instability of their
also characteristically had a greatly reduced tubulin MTs. Because of its close proximity to the taxane
polymer mass which is indicative of reduced MT binding site, this mutation may have an effect on
stability. The A231T substitution is located on H7 drug binding, although this is not likely.
within the highly hydrophobic region of the taxane
binding site (Figure 5a). Substitution of a hydro-
phobic alanine to a polar threonine residue should Mutations within the colchcicine
produce a significant effect on drug binding. binding site
However, as there was no observable effect on L240. The L240I mutation was identifi ed in tumor
paclitaxel binding, the authors suggested that the cell lines that became resistant to the MT desta-
change was more likely to result in the alteration of bilizing drug vincristine as a result of increased
the conformation of the H7, thereby affecting MT MT stability (Kavallaris et al. 2001). This muta-
stability through a reduction in the α helical propen- tion lies in a region of β-tubulin that is in close
sity associated with threonine over alanine.
proximity to the α /β-tubulin interface on the loop
Q292. Cells carrying a Q292E,H mutation were that links H7 and H8 (residues 238–250) (Figure 6a).
shown to become resistant to epothilone and A mutation at this position would affect longitu-
paclitaxel probably as a result of reduced drug dinal interactions between α and β monomers
binding (He et al. 2001; Verrills et al. 2003; Yang thereby stabilizing MTs conferring its observed
et al. 2005). This mutation was originally identifi ed vinca resistance phenotype. The authors proposed
as a suppressor of the mutation D45Y, originally that a leucine-to-isoleucine substitution at this
identifi ed by Wang et al. (2004). In the presence position is close to alanine 248 and may result in
of the mutation A231T, which was not shown to a conformational change within H7. As discussed
affect paclitaxel binding directly, mutations at previously, H7 lies at the base of the taxane
position 292 were shown to reduce paclitaxel binding site and has been implicated in the global
binding (Verills et al. 2003). As both of these muta-
stability of the tubulin dimer (Amos and Lowe,
tions resulted in reduced MT levels, the mutation 1999). This would support the observation that
of residue 292 may have a direct infl uence on cells carrying this mutation also had a concurrent
paclitaxel binding. Interrestingly, as can be seen increase in polymerized tubulin. Due to its
in Figure 5a Q292 is not in direct contact with the distance from the Vinca binding site, this mutation
bound paclitaxel molecule, but is located on H9 was hypothesized not to have an effect on drug
near the M loop, which is implicated in the critical binding, but is thought to lead only to increased
lateral contacts within the MT. This residue is also MT stability.
approximately 7.5 Å from the four-membered
D350. See the D197/K350 discussion below.
oxetane ring within paclitaxel or the ketone oxygen
within epothilone and may therefore affect the
binding pocket indirectly. Additionally, both gluta-
Mutations within the Vinca
mine and glutamic acids prefer solvent exposed binding site
conformations, they are frequently observed at S172. A single mutation at position S172 resulted
protein interfaces. The resulting change from in an overall increase in MT stability and a
glutamine to glutamic acid introduces a negative resulting resistance to the hemiasterlin HTI-286
charge which may decrease protofi lament-proto-
(Poruchynsky et al. 2004). This mutation also
fi lament interactions at this surface. This hypoth-
produced cross-resistance to vinblastine binding
esis is supported by the observation that cells site agents, as the hemiasterlins have been
carrying this mutation are sensitive to treatment proposed to share a binding region on tubulin with
with MT destabilizing drugs such as vinblastine. the vinca alkaloids (Bai et al. 1999). This mutation
When the Q292E was expressed in combination was shown to affect polymerization through the
with V60P cells became dependent on epothilone stabilization of MT thereby producing resistance
for growth (Yang et al. 2005). Both mutations at to destabilizing and sensitivity to polymerizing
Q292 and V60 would be expected to impact lateral drugs. The main reasoning for this may be due to
contacts between protofi laments. Therefore, cells the fact that S172 has recently been shown to be
carrying these mutations would require MT stabi-
a substrate for phosphorylation by cyclin-depen-
Cancer Informatics 2007: 3
The Roles of β-Tubulin Mutations and Isotype Expression in Acquired Drug Resistance
dent kinase Cdk1 at the transition from interphase close to the site where colcemid binds to tubulin
to mitosis (Fourest-Lieuvin et al. 2006). The and it is, therefore, unlikely to act by changing the
phosphorylation of this Serine residue was shown affi nity of tubulin for the drug. Wang et al. (2004)
to decrease MT stability and its loss would effec-
also identifi ed several revertants of D45Y MT
tively stabilize MTs, thereby making them more stabilization effects. V60 is also located in the H1-
resistant to destabilizing drugs. Therefore, resis-
S2 loop and its replacement with alanine results in
tance may be due an increase in stability or may the reduction of MT assembly rates, thereby
also be a result of reduced drug binding due to its conferring resistance to paclitaxel. These authors
proximity to the Vinca binding site.
observed that when D45Y and V60A substitutions
Interestingly, mutation of P173A has the oppo-
are co-expressed, their individual effects on MT
site effect leading to MT destabilization and resis-
assembly result in a normal phenotype. Because
tance to epothilone and sensitivity to colchicine both of these substitutions are found at a critical
(He et al. 2001). This difference may be due to a location for protofilament assembly, it is not
catastrophic conformational change in the T5 loop surprising that they display such a marked effect
as a result of the loss of P173. Neither of these on MT stability. An additional suppressor of the
mutations was shown to have any effect on drug D45Y phenotype, Q292 was also identifi ed by
Wang et al. (2004) to confer resistance to pacli-
P173. Mutations at P172 were shown not to taxel. This mutation resulted in the severe desta-
affect the binding of Epothilone and have a desta-
bilization of MTs and is located in H9, which is
bilizing effect on MT structure in HeLa cells (He positioned near the M-loop which is involved in
et al. 2001). P173 is located on the T5 loop which, MT lateral interface interactions and the taxane
as just described, is involved in forming the ribose binding site (Figure 5b). This position was also
binding component of the nucleotide binding shown to become mutated within human tumor
pocket (Figure 6b). It is highly probable that P173 cell lines that became resistance to desoxyepothi-
acts to stabilize this loop and is, therefore, essential lone B and epothilone B (Verrills et al. 2003; Yang
for proper nucleotide orientation within the nucle-
et al. 2005).
otide binding pocket. This would then have a direct
V60. As discussed above, mutations at V60
effect on nucleotide binding and subsequent hydro-
were observed under two separate studies which-
lysis, resulting in altered MT dynamics. Interest-
both resulted in decreased MT stability and reduced
ingly, while this mutation occurs next to S172, it assembly rates and a requirement for paclitaxel or
has the completely opposite effect, resulting in the epothilone presence for growth (Wang et al. 2004;
destabilization of MTs and a resistance to MT Yang et al. 2005). This mutation was originally
stabilizing drugs.
identifi ed as a suppressor of D45Y. Residue V60
is located at the end of the H1-S2 loop that has
been implicated as a principal partner of the
Mutations outside the drug
M loop for contacts between protofilaments
(Figure 5b). A mutation at residue 60 could poten-
D45. The mutant D45Y was shown to confer tially inhibit the lateral contacts between protofi la-
growth dependence on colcemid as a result of MT ments, resulting in the destabilization of MTs and
stabilization (Wang et al. 2004). This mutation is not predicted to confer altered drug binding.
caused an increase in the levels of polymerized
D197N/K350N. 2-Methoxyestradiol is a potent
tubuiln found within the cell, supporting the stabi-
anticancer agent that is thought to result in the
lization of MTs. This increase in stability could, destabilization of MT dynamics through its binding
therefore, explain the resistance to drugs such as at the colchicine binding site (D'Amato et al.
colchicine and vinblastine which destabilize MTs 1994). Cells expressing both the D197N and
and the increased sensitivity to those drugs like K350N mutations simultaneously became resistant
paclitaxel and epothilone that cause MT stabiliza-
to 2-Methoxyestradiol, colchicine and vinca
tion. This is most likely a result of D45 location alkaloids (Gokmen-Polar et al. 2005). This obser-
within a loop connecting helix 1 (H1) with β-sheet vation was suggestive of a global stabilization of
2 (S2), which was predicted to be a critical contact the MT structure with no effect on drug binding,
between tubulin dimers within protofilaments due to the different binding sites for vinblastine
(Figure 5b) (Li et al. 2002). This position is not and colchicine. Additionally, while these cells were
Cancer Informatics 2007: 3
resistant to MT destabilizing drugs, they did not
Y442. Identifi ed as a heterozygous mutation,
appear to be sensitive to MT stabilizing drugs such most likely due to a lethal phenotype in the homo-
zygous case. Substitution of Y442C resulted in
Several interactions for K350 and D197 illus-
cells that were dependent on either paclitaxel or
trate a possible structural basis of the resistance epothilone for growth (He et al. 2001). This obser-
to 2-methoxyestradiol when either is mutated to vation is, once again, suggestive of a global change
asparagine. First, K350 is not only located near in MT stability and not simply a decrease in overall
the colchicine binding site, but it also makes drug binding. This substitution occurs within the
critical longitudinal contacts with α tubulin as it end of H12 in a region not resolved in the crystal
is buried at the intra-dimer interface (Figure 6a). structure of the α/β-tubulin heterodimer (Lowe et al.
Large differences in IC50 values for both colchi-
2001). It is generally accepted that this region
cine and vinblastine binding also suggested that makes up the most variable part of the tubulin
drug binding was not affected as these two isotype sequences and is commonly referred to as
compounds bind at different sites within β-tubulin the C-terminal tail. This substitution is therefore
(Gigant et al. 2005). K350 may play a minor role most likely associated with the alterations of inter-
in stabilizing the α-phosphate moiety of GTP actions with MAPs or proteins involved in the post
bound tubulin, as it is only 6–7 Angstroms away. translational modification of the C-termini
Because it is in close proximity to the colchicine (Luduena, 1998).
binding site, K350's participation in direct interac-
tions with colchicine cannot be ruled out
Correlation Between β-Tubulin
Unlike K350, which forms bonds with portions Isotype Expression and Mutations
of α-tubulin, all of the local interactions for D197 We have examined the correlation between the
reside within β tubulin itself. D197, which resides positions of sequence differences associated
at the end of S6, participates in a series of hydrogen with β-tubulin isotypes and the mutations
bonds that are identical in both the straight and described here. Of all 21 mutations, only four
curved conformations of tubulin (Figure 5b) (Wang seem to correlate with residue differences
and Nogales, 2005). None of these hydrogen bonds between β-tubulin isotypes (Table 2, Figure 7).
would be signifi cantly disrupted by the mutation The first, at residue 45, is a conservative substi-
of D197N. However, D197 does participate in a tution from aspartic to glutamic acid and would
salt bridge with R156 within H4. Removal of this most likely not affect MT dynamics in the class
salt bridge would most likely lead to a loss of βII-VIII, isotypes that carry it. Next within the
anchoring of H4 and alter the propagation of human isotypes at position 172, we observed
conformational changes within β-tubulin, thereby the presence of a normal serine residue in class
infl uencing MT stability.
βI-VI and βVIII isotypes. However, in class
R306. Substitutions at position R306 were βVII this serine has been replaced by a leucine.
identifi ed in cells from paclitaxel resistant tumors As described above, this substitution results in
(Hasegawa et al. 2002). While this substitution did the loss of a polar serine. As this residue has
result in resistance to paclitaxel, there was no been shown to be phosphorylated during cell
determination if this was due to the destabilization division, its loss would lead to the resulting
of the MT structure, or a direct impact on drug stabilization of MTs into which it was incorpo-
binding. However, this substitution lies within the rated (Fourest-Lieuvin et al. 2006). While the
longitudinal interface between protofi laments and expression of class βVII has yet to be correlated
interacts with residue D118 within the adjacent with resistance to MT destabilizing compounds
β-tubulin monomer (Figure 5b). This interaction or increased MT stability, this is an avenue of
could constitute an additional salt bridge that acts experimentation worthy of future investigation.
to stabilize the longitudinal interface between The mutant A231T was shown to confer pacli-
protofi laments. Substitution to threonine would taxel or epothilone resistance onto cells that
result in the destruction of this interaction and expressed it (Verrills et al. 2003). Cells
could very possibly lead to an overall decrease in expressing this mutation were also shown to
MT stability, thereby producing a mechanism of exhibit greater levels of the class βIII isotype
resistance to paclitaxel.
expression. As previously stated, the A231T
Cancer Informatics 2007: 3
The Roles of β-Tubulin Mutations and Isotype Expression in Acquired Drug Resistance
Table 2. Hot spots correlating between β-tubulin isotype sequences and β-tubulin mutations associated with
acquired drug resistance. All residues identifi ed as being nonconserved among the Human β-Tubulin isotypes
were compared with those identifi ed in Table 1. The isotypes in which the change occurs are identifi ed with the
differing residues in parentheses.
Residue Mutant
Phenotype
β-tubulin Isotype Expression
βI(D) βII-VIII(E)
resistant βI-VI,VIII(S) βVII(L)β
resistant βI-V,VII,VIII(A) βVI(L)
resistant βI,VII(V) βIII,V(S) βII,IV,VI,VIII(A)
substitution is located on H7 within a highly the correlation between an isotype sequence and
hydrophobic region of the taxane binding site. an observed mutation is the presence of an
In the case of class βVI isotype containing a A364S substitution within the class βIII and βV
leucine at position 231, we expect that this isotypes. A364T was shown to become resistant
conservative substitution would not lead to a to paclitaxel but not epothilone (Giannakakou
dramatic change in phenotype.
et al. 2000). This substitution occurs within the
Finally, we feel that one of the most signifi-
taxane binding site and makes up a portion of
cant observations we have made with regard to the hydrophobic floor upon which paclitaxel
Correlation Between β-Tubuiln Isotypes and Mutations
Figure 7. Location of mutations and nonconserved residues in mammalian β-tubulin. As in Figure 3, with the exception that the β-Tubulin
surface has been removed to expose the underlying tertiary structure which is shown as a cyan cartoon. Here residues (F45, S172, A231 and A364), which are observed as occurring in both human β-tubulin isotypes and mutations identifi ed in Table 1, are shown as red sticks
in relation to paclitaxel (purple).
Cancer Informatics 2007: 3
and docetaxel's C13 side chain sits. The substi-
frequently exhibit increased sensitivity to drugs
tution of A364S in either class βIII or βV such as colchicine and vinblastine that bind to
isotypes would result in a similar situation to distinctly different sites. Instead of altered drug
that for the A364T mutant, a polar residue in binding, we favor a mechanism in which paclitaxel
this normally hydrophobic surface. This obser-
binding alters the conformation of both the M-loop
vation is exciting as the A364T mutation alone and the short loop connecting H6 and H7 of
was shown to impart selective paclitaxel resis-
β-tubulin in such a way as to facilitate and stabilize
tance. This, coupled with the observation that protein-protein interactions important in the forma-
the expression of the class βIII tubulin isotype tion of MTs. Recent fi tting of the crystal structure
may be correlated with paclitaxel resistance, of tubulin to a lower resolution map of the MT is
points to a possible mechanism and target for consistent with the participation of the H6-H7 loop
this resistance (Mozzetti et al. 2005).
in both longitudinal and lateral contacts (Amos and
Lowe, 1999). Because the mutations appear to
destabilize MT assembly in the absence of any drug
Even with the vast quantity of data available to us, a direct effect on protein-protein interactions
it is still not entirely clear what effect the expres-
appears the more likely possibility.
sion of a particular β-tubulin isotype, or acquisition
This hypothesis is also supported by the recent
of point mutations has on the stability of cellular observations of Xiao et al. (2006) who suggest
MTs and their relationship to drug resistance. Two that paclitaxel-binding may act to increase the
possible models may explain the role that these energy associated with lateral and longitudinal
two processes play in drug resistance. First, it may interactions between tubulin dimers in the MT.
be possible that certain isotypes or mutations Therefore changes within these surfaces would
exhibit decreased drug binding to β-tubulin. mitigate the action of energetic contributions of
However, our previous observations suggest that paclitaxel binding. Additionally, many of the
while this may contribute to drug resistance, it most mutations discussed here are not located within
likely plays a minimal role due to the lack of drug binding pockets at all, yet still confer drug
observable differences at the drug binding sites resistance onto the cells in which they are found.
(Huzil et al. 2006).
In general we can consider that cells resistant to
A second mechanism would produce global MT stabilizing drugs, such as the taxanes or
conformational differences between β-isotypes or epothilones, will produce less stable MTs in
mutants, thereby attenuating MT dynamics, response. This is demonstrated by decreased drug
making them more or less stable. We feel that the induced MT stability and often a requirement on
most interesting observation that has come out of the drug for growth. Conversely, cells that become
this examination of mutations within β-tubulin is resistant to MT destabilizing drugs such as colchi-
that out of a total of fi fteen residues that are found cine or vinblastine will produce MTs with
within the binding sites for either taxanes, colchi-
increased stability. We must, however, be cautious
cine or vinca alkaloids, only fi ve were defi nitively when examining the data available in the literature,
identified as having a direct impact on drug as much of the experimental results have been
binding. Although the mutations we have identifi ed obtained through in vivo screening and do not give
are close to the region implicated in the binding of us specifi c information about paclitaxel binding
paclitaxel, a number of observations indicate that to β tubulin. We believe that the key to developing
altered paclitaxel binding is not responsible for new drugs that target tubulin and disrupt MT
drug resistance in these mutants. For example, β- dynamics is through a better understanding of MT
tubulin is reported to contain the taxane binding dynamics at the molecular level and the role muta-
site, yet mutations conferring paclitaxel resistance tions and isotype differences have on the dynamic
occur with equal frequency in both α and β-tubulin properties of tubulin and the MT.
and both groups of mutants exhibit similar proper-
While it is clear that a substantial amount of work
ties (Minotti et al. 1991; Hari et al. 2003; Yang has to be done on obtaining specifi c binding data
et al. 2005). Many paclitaxel-resistant mutants for drug binding to β-tubulin isoforms and mutants
require the presence of the drug for cell division described here, the presence of minor variations
and, therefore, must clearly retain the ability to within the structure of β-tubulin may provide us
bind the drug. Also, paclitaxel-resistant mutants with a starting point for the development of novel
Cancer Informatics 2007: 3
The Roles of β-Tubulin Mutations and Isotype Expression in Acquired Drug Resistance
drugs, or in the derivatization of existing drugs in Blade, K., Menick, D.R. and Cabral, F. 1999. Overexpression of class I,
order to gain in increased specifi city or effect. This
II or IVb beta-tubulin isotypes in CHO cells is insuffi cient to con-fer resistance to paclitaxel. J. Cell. Sci., 112:2213–2221.
type of approach may also allow us to develop Boggs, B. and Cabral, F. 1987. Mutations affecting assembly and stabil-
secondary treatments for cancer cell lines that have
ity of tubulin: evidence for a nonessential beta-tubulin in CHO cells.
developed drug resistance as a result of standard
Mol. Cell. Biol., 7:2700–2707.
chemotherapy treatments, due to mutations or Bollag, D.M. et al. 1995. Epothilones, a new class of microtubule-stabiliz-
ing agents with a taxol-like mechanism of action. Cancer Res.,
altered expression levels. We feel that the most
signifi cant mechanism leading to drug resistance, Burkhart, C.A., Kavallaris, M. and Band Horwitz, S. 2001. The role of
as a result of both isotype expression and acquired
beta-tubulin isotypes in resistance to antimitotic drugs. Biochim. Biophys. Acta., 1471:O1–9.
mutations has to do with interactions within the MT Cabral, F., Abraham, I. and Gottesman, M.M. 1982. Revertants of a Chi-
structure itself and less to do with altered drug
nese hamster ovary cell mutant with an altered beta-tubulin: evi-
binding. This observation provides us with a novel
dence that the altered tubulin confers both colcemid resistance and temperature sensitivity on the cell. Mol. Cell. Biol., 2:720–729.
set of targets with which to begin designing drugs Cabral, F. and Barlow, S.B. 1989. Mechanisms by which mammalian cells
that target these "hot spots" in an effort to counter
acquire resistance to drugs that affect microtubule assembly. FASEB.
the effects that β-tubulin isotype expression or muta-
tions have on the development of drug resistance in Cabral, F., Sobel, M.E. and Gottesman, M.M. 1980. CHO mutants resis-
tant to colchicine, colcemid or griseofulvin have an altered beta-
these cells. We hope that the analysis presented here
tubulin. Cell., 20:29–36.
will provide deeper insights and perhaps a new Carles, G. et al. 1999. Differentiation of human colon cancer cells
direction for the future development of drugs that
changes the expression of beta-tubulin isotypes and MAPs. Br. J. Cancer, 80:1162–1168.
target crucial regions within the MT. It is our ulti-
Cowan, N.J. and Dudley, L. 1983. Tubulin isotypes and the multigene
mate goal to develop a new generation of drugs that
tubulin families. Int. Rev. Cytol., 85:147–173.
will target variant tubulins and thereby help reduce D'Amato, R.J. et al. 1994. 2-Methoxyestradiol, an endogenous mammalian
metabolite, inhibits tubulin polymerization by interacting at the
the adverse side effects associated with their use.
colchicine site. Proc. Natl. Acad. Sci. U.S.A., 91:3964–3968.
de Castro, J. et al. 2003. New insights in beta-tubulin sequence analysis
in non-small cell lung cancer. Lung. Cancer, 41:41–48.
Derry, W.B. et al. 1997. Taxol differentially modulates the dynamics of
This project was funded by grants from NSERC
microtubules assembled from unfractionated and purifi ed beta-tu-
(Canada), the Allard Foundation, MITACS and
bulin isotypes. Biochemistry, 36, 3554–3562.
Dozier, J.H. et al. 2003. Beta class II tubulin predominates in normal and
through support from Oncovista, LLC of San
tumor breast tissues. Breast Cancer Res., 5:R157–69.
Antonio, Texas and Technology Innovations, LLC Drechsel, D.N. et al. 1992. Modulation of the dynamic instability of tubulin
of Rochester, New York.
assembly by the microtubule-associated protein tau. Mol. Biol. Cell., 3:1141–1154.
Dumontet, C. et al. 1996. Differential expression of tubulin isotypes dur-
ing the cell cycle. Cell. Motil. Cytoskeleton, 35:49–58.
Ferguson, R.E. et al. 2005. Resistance to the tubulin-binding agents in renal cell
Achiwa, H. et al. 2003. Analysis of beta-tubulin gene alteration in human
carcinoma: no mutations in the class I beta-tubulin gene but changes in
lung cancer cell lines. Cancer Lett., 201:211–216.
tubulin isotype protein expression. Clin. Cancer Res., 11:3439–3445.
Amos, L.A. and Lowe, J. 1999. How Taxol stabilises microtubule structure.
Fourest-Lieuvin, A. et al. 2006. Microtubule regulation in mitosis: tubu-
Chem. Biol., 6:R65–9.
lin phosphorylation by the cyclin-dependent kinase Cdk1. Mol.
Bai, R. 1999. Interactions of the sponge-derived antimitotic tripeptide
Biol. Cell., 17:1041–1050.
hemiasterlin with tubulin: Comparison with dolastatin 10 and
Giannakakou, P. et al. 2000. A common pharmacophore for epothilone and
cryptophycin 1. Biochemistry, 38:14302–14310.
taxanes: molecular basis for drug resistance conferred by tubulin mutations
Banerjee, A. and Kasmala, L.T. 1998. Differential assembly kinetics of
in human cancer cells. Proc. Natl. Acad. Sci. U.S.A., 97:2904–2909.
alpha-tubulin isoforms in the presence of paclitaxel. Biochem.
Giannakakou, P. et al. 1997. Paclitaxel-resistant human ovarian cancer
Biophys. Res. Commun., 245:349–351.
cells have mutant beta-tubulins that exhibit impaired paclitaxel-
Banerjee, A. and Luduena, R.F. 1992. Kinetics of colchicine binding to
driven polymerization. J. Biol. Chem., 272:17118–17125.
purifi ed beta-tubulin isotypes from bovine brain. J. Biol. Chem.,
Gigant, B. et al. 2000. The 4 A X-ray structure of a tubulin:stathmin-like
domain complex. Cell, 102:809–816.
Banerjee, A. et al. 1990. Increased microtubule assembly in bovine brain
Gigant, B. et al. 2005. Structural basis for the regulation of tubulin by
tubulin lacking the type III isotype of beta-tubulin. J. Biol. Chem.,
vinblastine. Nature, 435:519–522.
Gokmen-Polar, Y. et al. 2005. beta-Tubulin mutations are associated with
Barlow, S.B., Gonzalez-Garay, M.L. and Cabral, F. 2002. Paclitaxel-de-
resistance to 2-methoxyestradiol in MDA-MB-435 cancer cells.
pendent mutants have severely reduced microtubule assembly and
Cancer Res., 65:9406–9414.
reduced tubulin synthesis. J. Cell. Sci., 115:3469–3478.
Gonzalez-Garay, M.L. and Cabral, F. 1995. Overexpression of an epitope-tagged
Bayley, P.M., Schilstra, M.J. and Martin, S.R. 1990. Microtubule dy-
beta-tubulin in Chinese hamster ovary cells causes an increase in endog-
namic instability: numerical simulation of microtubule transition
enous alpha-tubulin synthesis. Cell. Motil. Cytoskeleton, 31:259–272.
properties using a Lateral Cap model. J. Cell. Sci., 95:33–48.
Gonzalez-Garay, M.L. and Cabral, F. 1996. alpha-Tubulin limits its own
Berrieman, H.K., Lind, M.J. and Cawkwell, L. 2004. Do beta-tubulin mutations
synthesis: evidence for a mechanism involving translational repres-
have a role in resistance to chemotherapy? Lancet. Oncol., 5:158–164.
sion. J. Cell. Biol., 135:1525–1534.
Cancer Informatics 2007: 3
Gonzalez-Garay, M.L. et al. 1999. A beta-tubulin leucine cluster involved
Lowe, J. et al. 2001. Refi ned structure of alpha beta-tubulin at 3.5 A resolu-
in microtubule assembly and paclitaxel resistance. J. Biol. Chem.,
tion. J. Mol. Biol., 313:1045–1057.
Lu, Q. and Luduena, R.F. 1994. In vitro analysis of microtubule assembly
Gupta, M.L.J. et al. 2001. Mutagenesis of beta-tubulin cysteine residues in
of isotypically pure tubulin dimers. Intrinsic differences in the as-
Saccharomyces cerevisiae: mutation of cysteine 354 results in cold-
sembly properties of alpha beta II, alpha beta III, and alpha beta IV
stable microtubules. Cell. Motil. Cytoskeleton, 49:67–77.
tubulin dimers in the absence of microtubule-associated proteins. J.
Haber, M. et al. 1995. Altered expression of M beta 2, the class II beta-tu-
Biol. Chem., 269:2041–2047.
bulin isotype, in a murine J774.2 cell line with a high level of taxol
Luduena, R.F. 1998. Multiple forms of tubulin: different gene products and
resistance. J. Biol. Chem., 270:31269–31275.
covalent modifi cations. Int. Rev. Cytol., 178:207–275.
Hari, M. et al. 2003b. Mutations in alpha- and beta-tubulin that stabilize
MacRae, T.H. 1997. Tubulin post-translational modifi cations—enzymes
microtubules and confer resistance to colcemid and vinblastine. Mol.
and their mechanisms of action. Eur. J. Biochem., 244:265–278.
Cancer Ther., 2:597–605.
Maeno, K. et al. 2003. Mutation of the class I beta-tubulin gene does not
Hari, M. et al. 2003a. Expression of class III beta-tubulin reduces microtubule
predict response to paclitaxel for breast cancer. Cancer Lett.,
assembly and confers resistance to paclitaxel. Cell. Motil.
Mandelkow, E.M. and Mandelkow, E. 1992. Microtubule oscillations. Cell.
Hasegawa, S. et al. 2002. Mutational analysis of the class I beta-tubulin gene
Motil. Cytoskeleton, 22:235–244.
in human breast cancer. Int. J. Cancer, 101:46–51.
Margolis, R.L. and Wilson, L. 1981. Microtubule treadmills-possible mo-
He, L., Yang, C.P. and Horwitz, S.B. 2001. Mutations in beta-tubulin map
lecular machinery. Nature, 293:705–711.
to domains involved in regulation of microtubule stability in epothi-
Melki, R. et al. 1989. Cold depolymerization of microtubules to double
lone-resistant cell lines. Mol. Cancer Ther., 1:3–10.
rings: geometric stabilization of assemblies. Biochemistry,
Horio, T. and Hotani, H. 1986. Visualization of the dynamic instability of
individual microtubules by dark-field microscopy. Nature,
Mesquita, B. et al. 2005. No signifi cant role for beta tubulin mutations and
mismatch repair defects in ovarian cancer resistance to paclitaxel/
Huzil, J.T., Luduena, R.F. and Tuszynski, J.A. 2006. Comparative modelling
cisplatin. BMC. Cancer, 5:101.
of human and beta; tubulin isotypes and implications for drug binding.
Minotti, A.M., Barlow, S.B. and Cabral, F. 1991. Resistance to antimitotic
drugs in Chinese hamster ovary cells correlates with changes in the
Hyams, J.S. and Stebbings, H. 1979. The mechanism of microtubule associ-
level of polymerized tubulin. J. Biol. Chem., 266:3987–3994.
ated cytoplasmic transport. Isolation and preliminary characterisation
Mitchison, T. and Kirschner, M. 1984. Dynamic instability of microtubule
of a microtubule transport system. Cell. Tissue. Res., 196:103–116.
growth. Nature, 312:237–242.
Jensen-Smith, H.C., Luduena, R.F. and Hallworth, R. 2003. Requirement
Monzo, M. et al. 1999 Paclitaxel resistance in non-small-cell lung cancer
for the beta I and beta IV tubulin isotypes in mammalian cilia. Cell.
associated with beta-tubulin gene mutations. J. Clin. Oncol.,
Motil. Cytoskeleton, 55:213–220.
Jordan, M. and Wilson, L. 2004. Mary Ann Jordan, Leslie Wilson. Nat. Rev.
Mozzetti, S. et al. 2005. Class III beta-tubulin overexpression is a prominent
mechanism of paclitaxel resistance in ovarian cancer patients. Clin.
Kamath, K. et al. 2005. β III-Tubulin Induces Paclitaxel Resistance in As-
Cancer Res., 11:298–305.
sociation with Reduced Effects on Microtubule Dynamic Instability.
Nettles, J.H. et al. 2004. The binding mode of epothilone A on alpha,beta-
J. Biol. Chem., 280:12902–12907.
tubulin by electron crystallography. Science, 305:866–869.
Katsetos, C.D., Herman, M.M. and Mork, S.J. 2003. Class III beta-tubulin
Nicoletti, M.I. et al. 2001. Expression of beta-tubulin isotypes in human
in human development and cancer. Cell. Motil. Cytoskeleton,
ovarian carcinoma xenografts and in a sub-panel of human cancer
cell lines from the NCI-Anticancer Drug Screen: correlation with
Kavallaris, M., Burkhart, C.A. and Horwitz, S.B. 1999. Antisense oligonucle-
sensitivity to microtubule active agents. Clin. Cancer Res., 7:2912–
otides to class III beta-tubulin sensitize drug-resistant cells to Taxol.
Br. J. Cancer, 80:1020–1025.
Nogales, E., Wolf, S.G. and Downing, K.H. 1998. Structure of the alpha beta
Kavallaris, M. et al. 1997. Taxol-resistant epithelial ovarian tumors are as-
tubulin dimer by electron crystallography. Nature, 391:199–203.
sociated with altered expression of specifi c beta-tubulin isotypes. J.
Nogales, E. et al. 1995. Structure of tubulin at 6.5 A and location of the taxol-
Clin. Invest., 100:1282–1293.
binding site. Nature, 375:424–427.
Kavallaris, M. et al. 2001. Multiple microtubule alterations are associated
Odde, D.J., Cassimeris, L. and Buettner, H.M. 1995. Kinetics of microtubule
with Vinca alkaloid resistance in human leukemia cells. Cancer Res.,
catastrophe assessed by probabilistic analysis. Biophys. J., 69:796–
Keates, R.A., Sarangi, F. and Ling, V. 1981. Structural and functional al-
Panda, D. et al. 1994. Microtubule dynamics in vitro are regulated by the
terations in microtubule protein from Chinese hamster ovary cell
tubulin isotype composition. Proc. Natl. Acad. Sci. U.S.A., 91:11358–
mutants. Proc. Natl. Acad. Sci. U.S.A., 78:5638–5642.
Kemphues, K.J. et al. 1982. The testis-specifi c beta-tubulin subunit in Dro-
Pichichero, M.E. and Avers, C.J. 1973. The evolution of cellular movement
sophila melanogaster has multiple functions in spermatogenesis. Cell.,
in eukaryotes: the role of microfi laments and microtubules. Subcell.
Keskin, O. et al. 2002. Relating molecular fl exibility to function: a case study
Poruchynsky, M.S. et al. 2004. Tumor cells resistant to a microtubule-depo-
of tubulin. Biophys. J., 83:663–680.
lymerizing hemiasterlin analogue, HTI-286, have mutations in alpha-
Klein, L.E. et al. 2005. The microtubule stabilizing agent discodermolide is a
or beta-tubulin and increased microtubule stability. Biochemistry,
potent inducer of accelerated cell senescence. Cell. Cycle., 4:501–507.
Kurz, J.C. and Williams, R.C.J. 1995. Microtubule-associated proteins and
Prasannan, L. et al. 2000. Identifi cation of beta-tubulin isoforms as tumor
the fl exibility of microtubules. Biochemistry, 34:13374–13380.
antigens in neuroblastoma. Clin. Cancer Res., 6:3949–3956.
Li, H. et al. 2002. Microtubule structure at 8 A resolution. Structure (Camb),
Ranganathan, S. et al. 1998. Altered beta-tubulin isotype expression in pa-
clitaxel-resistant human prostate carcinoma cells. Br. J. Cancer,
Ling, V. et al. 1979. Mutants of Chinese hamster ovary (CHO) cells with
altered colcemid-binding affi nity. Cell., 18:423–430.
Ranganathan, S. et al. 1996. Increase of beta(III)- and beta(IVa)-tubulin
Liu, B. et al. 2001. Mechanisms of taxotere-related drug resistance in pan-
isotopes in human prostate carcinoma cells as a result of estramustine
creatic carcinoma. J. Surg. Res., 99:179–186.
resistance. Cancer Res., 56:2584–2589.
Cancer Informatics 2007: 3
The Roles of β-Tubulin Mutations and Isotype Expression in Acquired Drug Resistance
Ravelli, R.B. et al. 2004. Insight into tubulin regulation from a complex
Unger, E., Bohm, K.J. and Vater, W. 1990. Structural diversity and dynam-
with colchicine and a stathmin-like domain. Nature, 428, 198–202.
ics of microtubules and polymorphic tubulin assemblies. Electron.
Richards, K.L. et al. 2000. Structure-function relationships in yeast tubulins.
Microsc. Rev., 3:355–395.
Mol. Biol. Cell., 11:1887–1903.
VanBuren, V., Odde, D.J. and Cassimeris, L. 2002. Estimates of lateral and
Ringel, I. and Horwitz, S.B. 1991. Studies with RP 56976 (taxotere): a
longitudinal bond energies within the microtubule lattice. Proc. Natl.
semisynthetic analogue of taxol. J. Natl. Cancer Inst., 83:288–291.
Acad. Sci., U.S.A., 99, 6035–6040.
Roach, M.C. et al. 1998. Preparation of a monoclonal antibody specifi c for
Verdier-Pinard, P. et al. 2003. Direct analysis of tubulin expression in can-
the class I isotype of beta-tubulin: the beta isotypes of tubulin differ
cer cell lines by electrospray ionization mass spectrometry. Biochem-
in their cellular distributions within human tissues. Cell. Motil. Cy-
Verrills, N.M. et al. 2003. Microtubule alterations and mutations induced
Rowinsky EK, et al. 1989. Phase I and pharmacodynamic study of taxol in
by desoxyepothilone B: implications for drug-target interactions.
refractory acute leukemias. Cancer Res., 49:4640–7.
Chem. Biol., 10:597–607.
Sale, S. et al. 2002. Conservation of the class I beta-tubulin gene in human
Walker, R.A., Pryer, N.K. and Salmon, E.D. 1991. Dilution of individual
populations and lack of mutations in lung cancers and paclitaxel-
microtubules observed in real time in vitro: evidence that cap size is
resistant ovarian cancers. Mol. Cancer Ther., 1:215–225.
small and independent of elongation rate. J. Cell. Biol., 114:73–81.
Sangrajrang, S. et al. 1998. Association of estramustine resistance in human
Wang, H.W. and Nogales, E. 2005. Nucleotide-dependent bending fl exibil-
prostatic carcinoma cells with modifi ed patterns of tubulin expression.
ity of tubulin regulates microtubule assembly. Nature, 435:911–
Biochem. Pharmacol., 55:325–331.
Schiff, P.B. and Horwitz, S.B. 1980. Taxol stabilizes microtubules in mouse
Wang, Y., Veeraraghavan, S. and Cabral, F. 2004. Intra-allelic suppression
fi broblast cells. Proc. Natl. Acad. Sci. U.S.A., 77:1561–1565.
of a mutation that stabilizes microtubules and confers resistance to
Scott, C.A. et al. 1990. Beta-tubulin epitope expression in normal and
malignant epithelial cells. Arch. Otolaryngol Head. Neck. Surg.,
Wani, M.C. et al. 1971. Plant antitumor agents. VI. The isolation and struc-
ture of taxol, a novel antileukemic and antitumor agent from Taxus
Shalli, K. et al. 2005. Alterations of beta-tubulin isotypes in breast cancer
brevifolia. J. Am. Chem. Soc., 93:2325–2327.
cells resistant to docetaxel. FASEB. J., 19:1299–1301.
Weisenberg, R.C., Deery, W.J. and Dickinson, P.J. 1976. Tubulin-nucleotide
Soucek, K. et al. 2006. Normal and prostate cancer cells display distinct
interactions during the polymerization and depolymerization of
molecular profi les of alpha-tubulin posttranslational modifi cations.
Westermann, S. et al. 2006. The Dam1 kinetochore ring complex moves
Spiegelman, B.M., Penningroth, S.M. and Kirschner, M.W. 1977. Turnover
processively on depolymerizing microtubule ends. Nature, 440:565–
of tubulin and the N site GTP in Chinese hamster ovary cells. Cell.,
Westermann, S. and Weber, K. 2003. Post-translational modifi cations regu-
Sullivan, K.F. and Cleveland, D.W. 1986. Identifi cation of conserved iso-
late microtubule function. Nat. Rev. Mol. Cell. Biol., 4:938–947.
type-defi ning variable region sequences for four vertebrate beta tu-
Xiao, H. et al. 2006. Insights into the mechanism of microtubule stabiliza-
bulin polypeptide classes. Proc. Natl. Acad. Sci. U.S.A., 83:4327–
tion by Taxol. Proc. Natl. Acad. Sci. U.S.A., 103:10166–10173.
Yang, C.P. et al. 2005. A highly epothilone B-resistant A549 cell line with
Theodorakis, N.G. and Cleveland, D.W. 1992. Physical evidence for co-
mutations in tubulin that confer drug dependence. Mol. Cancer Ther.,
translational regulation of beta-tubulin mRNA degradation. Mol. Cell.
Zheng, Y. et al. 1995. Nucleation of microtubule assembly by a gamma-
Thompson, J.D., Higgins, D.G. and Gibson, T.J. 1994. CLUSTAL W: im-
tubulin-containing ring complex. Nature, 378:578–583.
proving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specifi c gap penalties and weight matrix choice. Nucleic. Acids. Res., 22:4673–4680.
Toso, R.J. et al. 1993. Kinetic stabilization of microtubule dynamic instabil-
ity in vitro by vinblastine. Biochemistry, 32:1285–1293.
Tsurutani, J. et al. 2002. Mutational analysis of the beta-tubulin gene in lung
cancer. Lung. Cancer, 35:11–16.
Cancer Informatics 2007: 3
Source: http://biomine.ece.ualberta.ca/papers/CINF-Tubulin2007.pdf
Im Auftrag der Stadtgemeinde Saalfelden seit 1996 Kinder & Jugendzentrum Saalfelden TREFFPUNKT · Berglandstraße 28 Gefördert durch die Stadtgemeinde Saalfelden und Mitteln des Land Salzburg Im Auftrag der Stadtgemeinde Saalfelden seit 1996 Im Wandel der Zeit! Das Kinder und Jugendzentrum Saalfelden „Treffpunkt", befi ndet sich ständig im Wandel. Viele der „älteren" Jugendlichen in der Altersgruppe von 15 bis 18 Jah-ren, haben nicht nur neue Interessen, sondern stehen auch vor neuen Herausfor-derungen wie dem Schulabschluss, den Beginn einer Lehre oder weiterführende Schule, der Führerscheinprüfung, dem Bundesheer oder Zivildienst und so wei-ter. Sie treffen sich im Jugendzentrum um sich diesbezüglich bei gleichaltrigen Alexander Houtman BEdauszutauschen und auch um Erfahrungswerte von BetreuerInnen anzuhören. Leiter des Kinder & Auffallend sind dabei die „kurzen" Besuche im Jugendzentrum - die Gespräche Jugendzentrum Saalfeldenmit den BetreuerInnen sind dafür aber intensiver.
Color profile: Disabled Composite Default screen Neisseria gonorrhoeae isolates in Metropolitan Toronto: Issues of scale, positional accuracy and confidentiality Jody F Decker PhD, Bob Sharpe PhD, Jo-Anne R Dillon PhD JF Decker, B Sharpe, J-AR Dillon. Mapping antibiotic-resistant Neisseria gonorrhoeae isolates in Metropolitan To-ronto: Issues of scale, positional accuracy and confidentiality. Can J Infect Dis 1997;8(5):273-278. The primary ob-jective of this paper was to investigate the methodological implications of mapping Neisseria gonorrhoeae using thepartial three-digit postal code instead of the complete six-digit postal code. The reporting locations of N gonorrhoeae iso-lates submitted from hospitals, doctor's offices, private and provincial laboratories, and sexually transmitted disease(STD) clinics were used as a model. Specifically, the paper focused on variations in geographical distributions of STDdata when mapped at different aggregations of postal code data and at different map scales. Such variations are of im-portance to those who analyze the spatial epidemiology of STDs, and the accessibility and use of health care services.This analysis showed that three-digit postal codes are useful in summarizing overall geographic distributions, butgreatly reduce positional accuracy, which can lead to inaccurate delineation of service areas and populations served. Thesix-digit postal code is more appropriate for detailed analysis addressing behavioural issues. The analysis demonstratedthat six-digits can be used without breaching individual confidentiality.