Biological effects of space radiation and development of effective countermeasures



Contents lists available at
Life Sciences in Space Research
Biological effects of space radiation and development of effectivecountermeasures
Department of Radiation Oncology, Perelman School of Medicine, University of Pennsylvania, Philadelphia, PA 19104-6072, United States
Article history:
As part of a program to assess the adverse biological effects expected from astronauts' exposure to space
Received 21 January 2014
radiation, numerous different biological effects relating to astronauts' health have been evaluated. There
Received in revised form 6 February 2014
has been major focus recently on the assessment of risks related to exposure to solar particle event (SPE)
Accepted 6 February 2014
radiation. The effects related to various types of space radiation exposure that have been evaluatedare: gene expression changes (primarily associated with programmed cell death and extracellular
matrix (ECM) remodeling), oxidative stress, gastrointestinal tract bacterial translocation and immune
Solar particle event
system activation, peripheral hematopoietic cell counts, emesis, blood coagulation, skin, behavior/fatigue
(including social exploration, submaximal exercise treadmill and spontaneous locomotor activity),
Acute effects of radiation
heart functions, alterations in biological endpoints related to astronauts' vision problems (lumbar
Long-term effects of radiation
puncture/intracranial pressure, ocular ultrasound and histopathology studies), and survival, as well aslong-term effects such as cancer and cataract development. A number of different countermeasures havebeen identified that can potentially mitigate or prevent the adverse biological effects resulting fromexposure to space radiation.
2014 The Committee on Space Research (COSPAR). Published by Elsevier Ltd. All rights reserved.
Changes in hematopoietic cell counts after proton or conventional reference radiation exposures . . . . . . . . . . . . . .
Changes in hematopoietic cell counts in mice after irradiation . . . . . . . . . . . . . . . . . . . . . . .
Changes in peripheral leukocyte counts in ferrets after irradiation . . . . . . . . . . . . . . . . . . . . . .
Changes in peripheral leukocyte counts in Yucatan minipigs after irradiation . . . . . . . . . . . . . . . . . .
Summary of effects of SPE radiation on hematopoietic blood cell counts in ferrets, mice and pigs . . . . . . . . . . .
Effect of SPE-like radiation on gastrointestinal tract integrity . . . . . . . . . . . . . . . . . . . . . . . .
Effect of SPE radiation and hindlimb suspension on immune function measured by bacterial challenge . . . . . . . . .
Summary of the effects of SPE radiation on the immune system . . . . . . . . . . . . . . . . . . . . . . .
Abbreviations: ARS, acute radiation sickness; aPTT, activated partial thromboplastin time; BBI, Bowman–Birk inhibitor; BBIC, BBI concentrate; BFO, blood forming or-
gans; BK, bradykinin; CNS, central nerve system; CT, computed tomography; DCF, dichlorofluorescein; DIC, disseminated intravascular coagulation; DMF, dose modifyingfactor; DNA-PKcs, DNA-dependent protein kinases; DTH, delayed type hypersensitivity; ECM, extracellular matrix; EGb76, quercetin; eSPE, simulated electron SPE; EVA, extra-vehicular activity; GI, gastrointestinal; GCR, galactic cosmic rays; G-CSF, granulocyte colony-stimulating factor; HDR, high dose rate; HS, hindlimb suspension; HZE particles,highly energetic, heavy, charged particles; ICRP, International Commission of Radiation Protection; IFN-α, interferon-alpha; INR, the patient's ‘test' PT value divided by thelaboratory ‘normal' PT value, raised to the power of the International Sensitivity Index; ISS, International Space Station; LAD, left anterior descending; LBP, lipopolysaccha-ride binding protein; LD50, dose expected to kill 50% of the treated subjects; LDR, low dose rate; LET, linear energy transfer; LPS, lipopolysaccharide; MnSOD, manganesesuperoxide dismutase; NAC, N-acetyl cysteine; NASA, National Aeronautics and Space Administration; NCRP, National Council on Radiation Protection and Measurements; NK,Natural Killer; PAMP, pathogen associated molecular patterns; PBMNC, peripheral blood mononuclear cell; PBS, phosphate buffered saline; PHA, phytohemagglutinin; PMN,polymorphonuclear leukocyte; pSPE, simulated proton SPE; PT, prothrombin time; PWS, partial weight suspension; RBE, relative biological effectiveness; SCR, solar cosmicradiation; SEB, surrogate endpoint biomarker; SeM, L-selenomethionine; SOBP, spread out Bragg peak; SPE, solar particle event; SWT, SI–Wu–Tang; TAS, total antioxidantstatus; TBI, total body irradiation; TF, tissue factor; vWF, von Willenbrand factor; WBC, white blood cell.
2214-5524/ 2014 The Committee on Space Research (COSPAR). Published by Elsevier Ltd. All rights reserved.
A.R. Kennedy / Life Sciences in Space Research 1 (2014) 10–43
Effects of radiation on blood coagulation and the development of disseminated intravascular coagulation . . . . . . . . . . .
Increased intracranial pressure and effects potentially related to vision abnormalities evaluated in Yucatan mini-pigs . . . . . . .
Histopathology changes in the eyes of pigs exposed to simulated proton and electron SPE radiation which could be related tovision problems . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
Ocular ultrasound results of the eyes of the pigs exposed to SPE radiation . . . . . . . . . . . . . . . . . . .
Opening pressure in pigs exposed to proton and electron SPE radiation . . . . . . . . . . . . . . . . . . . .
Countermeasures and mitigation of space radiation damage . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
Radiation induced oxidative stress and antioxidants as countermeasures . . . . . . . . . . . . . . . . . . . . . . .
Antioxidant protection against radiation induced cell death and transformation in vitro . . . . . . . . . . . . . . . . . .
Antioxidant protection against space radiation induced mortality . . . . . . . . . . . . . . . . . . . . . . . . .
Antioxidant protection against space radiation induced cataracts . . . . . . . . . . . . . . . . . . . . . . . . . .
Antioxidant prevention of space radiation induced cancer . . . . . . . . . . . . . . . . . . . . . . . . . . . .
Mechanism(s) for antioxidants as radiation countermeasures . . . . . . . . . . . . . . . . . . . . . . . . . . .
Granulocyte colony-stimulating factors as countermeasures . . . . . . . . . . . . . . . . . . . . . . . . . . . .
SI–Wu–Tang or fructose as countermeasures to increase neutrophil counts in mice exposed to SPE or γ -ray radiation . . . . . .
Antibiotics as countermeasures for bacterial toxicity in mice exposed to SPE radiation and HS . . . . . . . . . . . . . . .
Countermeasures for radiation induced emesis in ferrets . . . . . . . . . . . . . . . . . . . . . . . . . . . .
Countermeasures for altered bleeding times in ferrets after SPE radiation exposure . . . . . . . . . . . . . . . . . . .
Corticosteroid as a countermeasure for radiation induced pneumonitis and pneumonopathy in pigs . . . . . . . . . . . . .
Mometasone as a countermeasure for SPE proton radiation induced skin lesions in pigs . . . . . . . . . . . . . . . . .
Transparent film dressing for protection against proton therapy induced skin reactions in humans . . . . . . . . . . . . .
Summary concerning the major effects of SPE and space radiation . . . . . . . . . . . . . . . . . . . . . . . . . . . .
Changes potentially related to the development of vision abnormalities . . . . . . . . . . . . . . . . . . . . . . .
Threshold radiation doses for statistically significant adverse biological effects from SPE-like radiation in vivo . . . . . . . . . .
Agents identified as countermeasures for space radiation induced adverse biological effects . . . . . . . . . . . . . . . . . .
posed predominately of protons, with a minor contribution fromhelium ions (∼10%) and an even smaller contribution from heavy
As reviewed by the pri-
ions and electrons (∼1%). SPEs are unpredictable, develop rapidly
mary components of radiation in interplanetary space are galactic
and usually last for no more than several hours, although some
cosmic rays (GCR) and solar cosmic radiation (SCR). GCR originates
SPEs may continue for several days. Since protons are the major
from outside of our Solar System and consists of 98% baryons
component of SPE radiation, ground-based SPE radiation research
and 2% electrons. The baryonic component consists of 87% pro-
is focused on the biological consequences of proton radiation at
tons (hydrogen nuclei), 12% alpha particles (helium nuclei) and
the appropriate energies, doses, and dose-rates expected during an
approximately 1% of heavier nuclei with atomic numbers up to
SPE. A large fraction of the protons during an SPE are in the range
92 (uranium). These heavier nuclei include highly energetic, heavy,
of around 50 MeV, but there are also varying levels of protons
charged particles known as HZE particles. Although 56Fe ions, as a
of higher energies characterizing each individual SPE
specific type of HZE particle, account for less than 1% of the GCR
particle fluxes, they contribute significantly to the total radiation
Exposure to space radiation may place astronauts at signifi-
dose received by individual cells exposed to GCR due to the fact
cant risk for acute radiation sickness (ARS), significant skin injury
that the dose to an individual cell is proportional to the square of
and numerous other biological effects resulting from exposure to
the particle's energy dependent effective charge
radiation from a major SPE, which normally includes some HZE
SCR consists of low energy solar wind particles that flow con-
particles, or combined SPE and GCR. Doses absorbed by tissues
stantly from the Sun and the highly energetic solar particle events
vary for different SPEs and model systems have been developed
(SPEs) that originate from magnetically disturbed regions of the
to calculate the radiation doses that could have been received by
Sun, which sporadically emit bursts of energetic charged parti-
astronauts during previous SPEs For instance, it
has been estimated that the August 1972 SPE could have delivered
A.R. Kennedy / Life Sciences in Space Research 1 (2014) 10–43
doses of approximately 2.69 Gy and 0.46 Gy to skin and blood
ation Exposure, Risk of Acute Radiation Syndromes Due to Solar
forming organs (BFO), respectively, in a spacecraft and 32 Gy and
Particle Events (SPEs), Risk of Degenerative Tissue or other Health
1.38 Gy to skin and BFO, respectively, during extra-vehicular ac-
Effects from Radiation Exposure, and Risk of Radiation Carcino-
tivity (EVA). Depending on the radiation dose, dose rate and qual-
genesis. The Degenerative Tissue Risks include adverse radiation
ity, exposure to radiation during space missions may immediately
biologic effects on the heart, circulatory, endocrine, digestive, lens
affect the probability for successful mission completion (mission
and other tissue systems (which would include radiation effects on
critical) or result in late radiation effects in individual astronauts
bone, muscle, etc.). It is noteworthy that the International Commis-
While avoidance of the radi-
sion of Radiation Protection (ICRP) has recently issued a report that
ation risk is the best protective strategy, it is nearly impossible
has important implications for two of the degenerative tissue risks,
to avoid the radiation risk completely for astronauts. Therefore,
circulatory diseases and cataracts In this re-
countermeasures against adverse biological effects of space radi-
cent review of early and late effects of radiation in normal tissues
ation are necessary for the success of long term space missions.
and organs, it was concluded that for reactions manifesting very
National Aeronautics and Space Administration (NASA) is primar-
late after low total doses, particularly for cataracts and circulatory
ily concerned with the health risks for astronaut exposures to GCR
disease, it appears that the rate of dose delivery does not mod-
and SPE radiation. SPEs occur with variable tissue dose-rates and
ify the incidence, and for these two tissues, a threshold dose of
doses, which range from 0 to 0.5 Gy/hour and 0 to 2 Gy, respec-
0.5 Gy was proposed For a NASA mission to
tively, and with skin doses > 5 Gy NASA has
Mars, fatal cancer risk has been considered the dominant risk in
determined that the likelihood of acute risks during internal vehi-
the past (considering the dose from GCR), but circulatory diseases
cle activity is extremely small; however, there are scenarios during
are likely to be of great importance in the newer risk estimates
lunar, trans-lunar or Mars EVAs in which ARS may occur.
for a mission to Mars There have been a
Acute radiation sickness has a sequence of a phased syndrome
number of recent reviews or updates on the status of space radi-
that varies with radiation dose, dose rate, quality and individual ra-
ation research in the research areas of particular concern to NASA
diation sensitivity which can
for the exploration class missions planned for the future.
include nausea, vomiting, diarrhea and fatigue. These effects are
For long term space exploration, bone loss and muscle atrophy
manifested at approximately 4 to 24 hours post-exposure for expo-
due to disuse are other major concerns related to the health of
sures at sub-lethal doses, with a latency time inversely correlated
the crew In ground-
with dose. Since exposure to proton radiation, which represents
based studies, disuse bone loss has been observed in the hindlimb
the major type of radiation in an SPE, is known to induce ab-
suspension rodent model
normalities in leukocytes, erythrocytes and platelets
there is also a reasonable concern for compromised
with γ -rays exacerbates skeletal microarchitectural changes that
immune functions, especially in the microgravity environment in
are normally found during progressive, postpubertal aging prior to
the onset of age-related osteoporosis Radi-
Space flight is known to alter immune
ation exposure may increase the number of osteoclasts and the
responses, and the causal factors include the stress due to in-
extent of acute bone loss via increased reactive oxygen species
creased radiation exposure
production and oxidative damage, which implies different molec-
and microgravity and non-load bearing sta-
ular mechanisms from the bone loss caused by disuse
Irradiation with 250 MeV protons followed by hindlimb
The consistent effects on the immune
suspension resulted in an approximately 20% loss of the trabec-
system observed so far during space travel are as follows: reduc-
ular bone volume fraction in the tibia and femur of irradiated
tion in peripheral T-cell counts and a decrease in Natural Killer
mice, and the mice receiving the combined treatment with proton
(NK) cell number and functionality
radiation and hindlimb suspension generally experienced greater
decreases in cell-mediated immunity
loss of the trabecular bone volume fraction, connectivity density,
with altered cytokine production
and trabecular number than either hindlimb suspension or irra-
but normal levels of serum immunoglobulins
diation treatment alone Irradiation with 56Fe
An increased susceptibility to infection under
ions, which represents a significant component of GCR
space flight conditions has also been observed
stimulates osteoclast differentiation even in the absence of
The main concern of an impaired immune
osteoblasts, thereby enhancing the sensitivity of bone cells to the
system in the closed environment of a spacecraft is the altered
effects of radiation Iron ion radiation con-
ability to control bacterial, fungal, viral, and parasitic invasions
tributes to a reduction in compressive strength and partially pre-
vents the recovery of cancellous microarchitecture from adaptive
and the loss of immuno-
responses of lumbar vertebrae to skeletal unloading in hindlimb
surveillance leading to tumor growth Counter-
suspended mice Thus, irradiation with heavy
measures that have been considered and/or evaluated for mitigat-
ions may accelerate or worsen the loss of skeletal integrity trig-
ing acute radiation effects on immune system include interferons,
gered by musculoskeletal disuse in the microgravity environment.
which have a profound effect on the immune response both in
There are some publications indicating that countermeasures may
vivo and in vitro an active hex-
be helpful to mitigate radiation induced adverse bone effects; for
ose correlated compound, which activates immune function
example, α-lipoic acid protects cancellous tissue from the detri-
and enhances resistance to infection
mental effects of irradiation
and vitamin and mineral dietary supplementation, as
The space radiation risks to the central nerve system (CNS)
recently reviewed
have been considered to be extremely important in the recent
In addition to acute effects from radiation, there are numerous
past due to major publications in this field of research. Examples
other major health concerns related to space radiation exposure.
of such publications include studies suggesting that the induction
In the NASA Human Research Roadmap (A Risk Reduction Strat-
of Alzheimer's disease may be a space radiation risk
egy for Human Space Exploration), the Integrated Research Plan
and attention deficits may arise following exposure to
(IRP) divides the space radiation risks into the following categories:
low doses of space radiation Many other
Risk of Acute and Late Central Nervous System Effects from Radi-
publications have also indicated that there are major CNS space
A.R. Kennedy / Life Sciences in Space Research 1 (2014) 10–43
radiation risks [e.g.,
ment of hepatocellular carcinoma There are
some intriguing recent results in the radiation carcinogenesis field
Radiation exposure to
of research. have indicated that there are distinct
γ -rays or 56Fe ions
signatures (transcriptome profiles) in normal human bronchial ep-
ithelial cells exposed to γ -rays and different HZE particles. If this
to have adverse effects on CNS and neurobehavior of irradiated an-
effect can be confirmed, it may give rise to studies in which the
imals, including reduced performance in motor tasks and deficits
causative agent can be identified in human malignancies that could
in spatial learning and memory. Alterations in neuronal function
have been caused by radiation exposure. While the mechanism(s)
in the HZE particle irradiated animals include reduced responsive-
involved in space radiation induced carcinogenesis are still un-
ness to agonist stimulation and increased Nigral cell loss, which
known, there is evidence that space radiation induced oxidative
parallel the neurobehavioral changes associated with aging
stress is closely associated with carcinogenesis [e.g.,
The available data suggest that: a) the neu-
It has been reported that space radiation induces per-
rochemical and behavioral deficits after HZE radiation exposure
sistent oxidative stress in mouse intestine, which is likely to be
have an apparent threshold below which there are no effects on
associated with intestinal tumorigenesis There
these endpoints, b) there does not appear to be a dose–response
is evidence that space radiation induced carcinogenesis can be
curve for many endpoints, such as upper body strength or radia-
prevented or mitigated by several cancer chemopreventive agents,
tion induced taste aversion learning, and c) there is no evidence
which include antioxidants and protease inhibitors [as recently
of spontaneous recovery of function that depends upon the in-
reviewed retinoids [e.g.
tegrity of the dopaminergic system after the HZE radiation ex-
and fruit extracts
posure It has been reported that persistent
In addition, there are a number of new potential
radiation induced oxidative stress is associated with space radi-
cancer preventive agents that have been shown to mitigate in vitro
ation induced CNS effects [e.g., In the CNS
SEBs of the space radiation cancer risk; examples include mela-
research area, there have been several publications indicating that
tonin and a synthetic triterpenoid, bardoxolone
countermeasures exist for some of the space radiation induced
methyl, which protects against space radiation-induced transfor-
adverse biological effects, which include melatonin or a metabo-
mation of human colon epithelial cells
lite lipoic acid
There is extensive evidence that radiation exposure on earth
fruit extracts, which ameliorate deficits in behavior
can give rise to cardiovascular diseases, as recently reviewed
and signaling in rats irradiated with 56Fe ions
and flavonoid glycosides from Ginkgo biloba, myricetin and
the research on heart and circulatory effects resulting from ex-
quercetin (EGb761) which have been pos-
posure to space radiation(s), it has recently been reported that
tulated to improve cerebral metabolism, protect the brain against
doses of 2 to 5 Gy 56Fe ion radiation targeted to specific arte-
hypoxic damage and scavenge free-radicals
rial sites in apolipoprotein E-deficient (apoE−/−) mice accelerated
Carcinogenesis has continued to be the major focus of the NASA
the development of atherosclerosis In these stud-
space radiation risk experimental investigations over the past sev-
ies, it was concluded that 56Fe ions can promote the progression
eral years, with most of the investigations not focused on the
of atherosclerotic lesions to an advanced stage characterized by
development of tumors in animals developing from space radia-
compositional changes indicative of increased thrombogenicity and
tion exposure(s), but instead focusing on various potential surro-
instability. In numerous other studies, space radiation has been
gate endpoint biomarkers (SEBs) of space radiation carcinogenesis.
shown to have detrimental effects on many other parameters re-
There are some recent reviews that focus on the development
lated to cardiovascular and circulatory diseases, with particularly
of cancer in animals exposed to space radiation [e.g.,
strong effects leading to endothelial dysfunction [e.g.,
and angiogenesis [e.g.,
some recent individual re-
ports on space radiation induced tumorigenesis [e.g.,
Risks of other degenerative diseases include radiation induced
cataracts, as recently reviewed
and some new hypotheses/thoughts concerning mechanisms of ra-
diation carcinogenesis [e.g.,
It is noteworthy that the
and risk estimates of radiation induced cancer [e.g.,
ICRP has recently proposed lowering the threshold for radiation
induced cataracts to 0.5 Gy There have
ing in the recent animal carcinogenesis studies is that 56Fe ions
also been some recent studies on space radiation (or other sim-
were not substantially more effective than γ -rays for the induc-
ilar types of radiation such as heavy ion-cancer therapy [hadron
tion of acute myeloid leukemia
therapy]) induced cataracts
It has been pointed out in numerous cur-
rent and older reviews of space radiation carcinogenesis stud-
has been reported that astronauts have an elevated risk of devel-
ies that space radiation induced malignancies are dependent on
oping cataracts which
the species as well as the strain of the species used, and that
has been associated with exposure to the high linear energy trans-
a major task in this field of research will involve determina-
fer (LET) GCR present in the space environment.
tions about the appropriate methods to use for extrapolation of
Current medical treatment for the acute radiation syndrome
the space radiation induced cancer risks from experimental ani-
routinely includes supportive care, antibiotics (quinolones and
mal studies to humans. One example of the differences observed
other agents), cytokine therapy, anti-emetic agents and analgesic
in space radiation induced cancer studies concerns the develop-
agents Other agents
ment of hepatocellular carcinoma. While exposure to space ra-
can also be used for the effects of the acute radiation syndrome,
diation(s) has indicated a very high incidence of hepatocellular
such as antihistamines, anti-inflammatories and radioprotectors
carcinoma in one mouse strain
Several FDA approved anti-emetic drugs, such
in other experiments on space radiation induced
as Kytril (granisetron), Zofran (Ondansetron), Decadron® (dexam-
carcinogenesis using a different strain of mice, a dose of 0.5 Gy
ethasone tablets) and Emend (Aprepitant) are known to prevent
from 56Fe ions or 3 Gy from protons had no effects on the develop-
or alleviate nausea and vomiting in patients or animals exposed
A.R. Kennedy / Life Sciences in Space Research 1 (2014) 10–43
to radiation or chemotherapeutic agents
of SPE radiation with conventional γ -ray radiation. The dose dis-
tribution from electron radiation, however, can be manipulated to
A systematic review and meta-analysis of 14 ran-
simulate the dose distribution expected from SPE radiation. Mega-
domized controlled trials, comprising 1451 patients, showed that
voltage electron beam radiation has been utilized in the pig exper-
amifostine (WR2721) significantly reduced the side effects of ra-
iments to accurately reproduce the total dose and dose distribution
diation therapy In the animal studies, treat-
of SPE protons The dosimetry involved in de-
ment with amifostine was shown to protect against DNA damage
termining simulated SPE radiation doses in pigs is illustrated in
in cisplatin treated murine peripheral leukocytes
which shows that the doses to the external organs (e.g.,
reduce changes in nucleolar morphology in-
skin, lens) are very high, while the doses to internal organs (e.g.,
duced by cisplatin treatment and protect
spinal cord, bone marrow) are quite low. The detailed methods for
against cyclophosphamide-induced disruption of taste
determining the organ doses are described elsewhere
and methotrexate-induced small intestinal mucosi-
These methods incorporate
tis as well as inhibit tumorigenesis
modern radiation oncology approaches of computed tomography
Unfortunately, the severe side effects of am-
(CT) based Monte Carlo dosimetry into the studies so that acute
ifostine have limited its use in the space program for astro-
biological effects in specific organ systems can be determined in
nauts as well as in other human populations exposed to ra-
animal model systems, and radiation toxicity from various types of
diation. PrC-210 is a new aminothiol that has shown no de-
SPE radiation exposures can be compared. This approach has also
tectable nausea/vomiting or hypotension side effects in the fer-
been used to predict the acute biological effects of SPE radiation
ret and rat models in contrast to the strong
exposure in astronauts. Ten full body human CT scans in various
side effects of the current aminothiol, amifostine
geometries have been analyzed to determine the impact of physical
this compound shows promise as a new
and environmental factors on organ dosimetry in humans. It has
been found that, depending on the organ system of interest (deep
In the past several years, we have been engaged in research
vs. superficial) and the fluence/energy profile of the exposure (hard
to assess whether there will be adverse acute biological effects
vs. soft event), either the physical size of the astronaut or the flu-
similar to those of ARS after exposure to the types of radiation
ence/energy profile for the SPE can be the determining factor for
at the energies, doses and dose-rates expected during an SPE.
radiation dose/toxicity (Cengel, K.A., Schaettler, M.O., and Diffend-
The overall objectives of our studies were to assess the risk of
erfer, E., Unpublished data). In contrast, most of the experiments
ARS and evaluate countermeasures for ARS, which can develop
with mice or ferrets utilizing SPE-like proton radiation involved
after exposure to SPE radiation. There is also a reasonable con-
homogeneous proton radiation exposures with relatively low en-
cern for a compromised immune system, due to high skin doses
ergies like those present in SPEs; RBE values were then calculated
from an SPE, which can lead to burns. Existing evidence sug-
by comparison of the SPE-like proton results with those obtained
gests that the best animal model for radiation induced vomit-
in similar experiments using γ -ray radiation.
ing is the ferret whereas the
In this review paper, the results of our studies on the bio-
best animal model for radiation induced skin changes is the pig
logical effects of several different types of space radiation, which
include different types of SPE radiation, are discussed. Both acute
Mice, on the other hand, are the most
and chronic effects of space radiation have been evaluated in these
frequently utilized mammalian species for evaluation of many ra-
diation induced biologic effects. A major problem concerning theuse of mice for studies of SPE radiation is that mice do not
2. Acute radiation effects
vomit in response to irradiation There-fore, three species of animals, i.e., ferrets, pigs and mice, were
The acute radiation effects evaluated in our studies include
used in our studies to allow interspecies comparisons of the re-
effects on hematopoietic cells, immune system effects (which in-
sults obtained in the studies in several different areas of research,
clude immune system changes resulting from a high dose of SPE
whereas the effects on vomiting and skin were evaluated only in
radiation to the skin), behavior/fatigue, heart functions, skin ef-
the most appropriate animal species. Since astronauts will be ex-
fects and organism survival after exposure to a lethal or potentially
posed to space radiation in a microgravity environment, which is
lethal dose of radiation.
known to cause bone loss, muscle atrophy and injury to soft con-nective tissues in animal models some of the radiation experiments with
2.1. Changes in hematopoietic cell counts after proton or conventional
mice have been performed with and without partial weight sus-
reference radiation exposures
pension (PWS) at one-sixth G, which is known to be the gravityon the surface of the Moon or hindlimb sus-
The changes in peripheral leukocytes in animals post-irradiation
pension (HS), a model appropriate for mimicking travel in deep
have been evaluated in ICR mice irradiated with 225-kVp X-rays
space to evaluate and quantify the
γ -rays from 60Co or
possible synergy between radiation and simulated hypogravity on
hematopoietic effects associated with ARS.
protons with energies of 50-MeV
In the studies in which SPE radiation effects have been evalu-
ated, it was considered extremely important to have comparable
dose distributions between the SPE radiation and a standard ref-
78.4 MeV and 1-GeV pro-
erence radiation in the animal model systems so that relative bio-
tons as well as pro-
logical effectiveness (RBE) values could be calculated
tons with eight different energies between 31 and 75 MeV and
SPE radiation is known to result in an inhomogeneous total
simulated SPE protons with energies between 30 and 150 MeV
body distribution, with a considerably higher dose delivered to the
The changes in peripheral leukocytes have
skin and underlying tissues than to the internal organs
also been examined in ferrets irradiated with 60Co or 137Cs
These characteristics of an SPE, which result in an unusual
γ -rays and 110-MeV protons
dose distribution pattern, make it difficult to compare the results
and Yucatan minipigs irradiated with 6-MeV electrons

A.R. Kennedy / Life Sciences in Space Research 1 (2014) 10–43
Fig. 1. Simulated dosimetry for Yucatan minipigs. Modern radiation oncology approaches, using CT based Monte Carlo dosimetry, have been incorporated into recent studies
to accurately predict specific organ doses from SPE radiation exposure in animals. Pig data are shown in the figures. A: 3D reconstruction from a pig CT image with the skin
rendered translucent to allow viewing of the internal organs (i.e., spinal cord [white], bone marrow [red], lung pleura [blue]). The organs were identified on separate CT axial
cross sections generating a 3D organ volume from the combined axial contours. (This figure has been reproduced with permission from Radiation Research B: Dose overlay on the 3D rendering of the whole body CT scan. (This figure has been reproduced with permission from Radiation Research C:
Dose overlay on a pig CT image (coronal plane cross-section) for 6 + 12 MeV electron irradiation (often called a Heat map or a Dose-Color map) (Doses were simulated for
6 + 12 MeV electron irradiation of a pig using a Monte Carlo based simulation algorithm (Varian Medical Systems, Palo Alto, CA). D: Dose–volume histogram (DVH figure)
illustrates organ radiation doses determined using the organ volumes and simulated dose distribution for the pigs receiving a skin dose of 20 Gy (from electrons). Note the
high skin dose in the dose–volume histogram (DVH) figure and the fact that the lens dose is also very high. (Images C and D – courtesy of Dr. Eric Diffenderfer.)
6 + 12-MeV electrons and SPE protons
were still 64% and 76% below the respective control values
whereas peripheral WBC and lymphocyte counts inthe mice irradiated with 5.9 Gy of 1-GeV protons were still ap-
2.1.1. Changes in hematopoietic cell counts in mice after irradiation
proximately 37% and 44% below the respective control values in
In an early study focused on determining the changes in circu-
non-irradiated control animals In contrast,
lating hematopoietic cell counts in mice exposed to SPE-like proton
the PMN/neutrophil count was fully recovered by 8 weeks post-
or γ -ray radiation (used as the reference radiation), ICR outbred
irradiation with 8 Gy of 225 kVp X-rays
mice aged 5–6 weeks were exposed to 60Co γ -rays at doses of
In a separate study performed with male ICR mice aged 4–5
0.13, 0.25, 0.5, 1 or 2 Gy or spread out Bragg peak (SOBP) protons
weeks, exposure to 1 GeV proton radiation administered in a sin-
(50 or 70 MeV) at doses of 0.25, 0.5, 1 and 2 Gy. The radiation
gle dose at low (5 cGy/minute) or high (50 cGy/minute) dose rates,
exposures were delivered in a single dose at the low dose rate
or in five fractionated doses at the low dose rate resulted in signif-
of 0.5 Gy/hour or the high dose rate of 0.5 Gy/minute. The re-
icant and dose dependent decreases in peripheral WBC and lym-
sults demonstrated a dose dependent decrease in white blood cell
phocyte counts at 24 hours post-irradiation
(WBC) counts in mice exposed to high and low dose rate proton
However, the difference among animals irradiated with the five
and γ -radiation
fractionated doses, or in a single dose at low or high dose rate, did
In 4–5 weeks old ICR mice, peripheral WBC, lymphocyte and/or
not reach statistical significance at any of the doses evaluated, and
polymorphonuclear leukocyte (PMN) counts decreased significantly
neither the WBC counts nor the lymphocyte counts in the animals
at 4 and 24 hours after total body irradiation with 1 or 8 Gy of
irradiated with 2 Gy 1-GeV protons in the five fractionated doses,
225 kVp X-rays or 1 or 5.9, 6.8 or 7.2 Gy of
or in a single dose at low or high dose rate, were significantly
1 GeV protons At 24 hours after irradiation
different from the animals irradiated with 2 Gy of 51.24-MeV pro-
with 8 Gy of 225 kVp X-rays, the neutrophil count was decreased
tons administered at a low dose rate (5–7 cGy/minute), although
to an average of 450 cells/μl; neutrophil counts of <500 cells/μl are
they were all significantly below the control WBC and lymphocyte
clinically significant. At 8 weeks post-irradiation, peripheral WBC
counts in sham irradiated animals. These results suggest that the
and lymphocyte counts in the mice irradiated with 8 Gy of X-rays
effect of proton irradiation on the WBC and lymphocyte counts in
A.R. Kennedy / Life Sciences in Space Research 1 (2014) 10–43
the irradiated mice is not altered by dose fractionation, dose rates
evaluate the impact of hypogravity on the effect of SPE radia-
or proton energy in the ranges evaluated
tion on immunological function, experiments were performed with
Some experiments were performed to determine whether age
6–8 weeks old female ICR mice that were irradiated with 0.5, 1
or sex/gender differences affected the ability of SPE-like radiation
or 2 Gy of γ -rays with or without hypogravity simulated using
to affect circulating hematopoietic cell counts in mice. To deter-
the PWS model, described by The combi-
mine the effects of age on this endpoint, 1-year old ICR mice
nation treatment with PWS and γ -ray irradiation decreased total
were exposed to proton or γ -ray radiation at doses of 0.0, 0.5,
splenic lymphocyte viability in a dose dependent manner, and the
1.0, or 2.0 Gy, with a dose rate of 0.5 Gy/min
suppressed splenic lymphocyte viability in groups exposed to a
Whole blood samples were collected
2 Gy dose of radiation persisted for 4 days, which was the last
up to 30 days post-irradiation and complete blood counts were an-
time point evaluated in the study In addi-
alyzed using automated technology as previously described
tion, the viability of splenic lymphocytes was significantly lower
in the mice that received a 1 Gy dose of γ -rays in combina-
A comparable experiment was performed in young
tion with PWS treatment than in the mice that received a 1 Gy
ICR mice, aged 6–8 weeks. Statistical analyses and RBE values were
dose of γ -rays without PWS treatment on Day-1 or Day 4 post-
determined as previously described
irradiation. Treatment with PWS alone did not significantly af-
for the experiments performed in both young and aged mice. The
fect the splenic lymphocyte viability at any of the time points
older mice in this experiment were specifically aged to simulate
evaluated up to 4 days. These results suggest that simulated hy-
healthy, non-smoking, middle-aged astronauts. For example, the
pogravity might have made splenic lymphocytes more sensitive
preferred age range for European Space Agency applicants is 27–37
to the cell killing effects of radiation. In addition, results from
years old. The decline in WBCs, neutrophils, lymphocytes and gran-
these studies indicated that T cell activation was decreased in the
ulocytes were not different using aged mice as compared to the
irradiated mice (1 or 2 Gy) with or without simulated hypograv-
reduction observed in young mice (Sanzari, J.K. and Kennedy, A.R.,
ity (PWS). Similar results were observed in experiments using
Unpublished data). The lymphocyte nadir was at 2 days post-
mice exposed to SPE-like proton radiation with and without sim-
proton irradiation, with counts as low as 50%, 60%, and 77% of
ulated hypogravity produced by HS
the control values after exposure to 0.5, 1.0 and 2.0 Gy protons, re-
In these experiments, mice were suspended
spectively. The decline in lymphocyte counts after γ -ray radiation
prior to and after SPE proton radiation exposure and total leuko-
exposure was similar to that observed after the proton radiation
cyte numbers and splenic lymphocyte functions were evaluated
exposure, which were also comparable to the decline observed in
on days 4 or 21 after the radiation exposure with and without
young mice. The results for the granulocyte counts were also con-
HS. Splenic lymphocyte subpopulations were altered at both time
sistent between the aged and young mice, with a bimodal decline
points investigated. At 21 days post-exposure, T cell activation
observed for the first nadir at 4 days post-irradiation (γ -ray or
and proliferation were assessed in isolated lymphocytes. In these
proton) and the second nadir at 16 days post-irradiation. The RBE
studies, T cell activation was suppressed in the proton-irradiated
values at different time points and for each dose were calculated
animals and in the irradiated animals exposed to HS. From both
in the aged and young mice. It was observed that the RBE val-
types of experiments described above, the results suggest that
ues were not significantly different from 1.0 in either the young
these irradiated animals with or without additional exposure to
or the aged mice. In similar experiments, the radiation effects on
simulated microgravity would have immune system suppression
WBCs, lymphocytes and granulocytes were shown to be similar be-
resulting from the lack of T cell activation. However, the peripheral
tween male and female CHH/HeN mice
blood cell (lymphocyte and granulocyte) counts were significantly
The only sex/gender difference observed in these experiments was
higher in proton irradiated mice with HU treatment than without
that non-irradiated male mice had 13% higher platelet counts and
HU treatment and the HU treatment did not significantly interact
more enhanced recovery of platelets on day 16 after irradiation as
with the proton radiation dose in the blood cell count data, indi-
compared to female mice. Thus, it is conceivable that this differ-
cating that the effects of radiation and hypogravity on peripheral
ence between male and female mice could influence the response
leukocytes were simply additive (or subtractive) with no significant
of platelets to total body radiation exposure.
In a study conducted with 5 to 7-weeks old female ICR mice,
To determine the RBE values for the effects of SPE protons in
exposure to protons with 8 energy levels ranging from 30.63 to
hematopoietic cells, several studies were performed in mice us-
74.52 MeV or simulated SPE protons with energies ranging from
ing 21-MeV electrons 60Co γ -rays
30 to 150 MeV at high (0.5 Gy/minute) or low (0.5 Gy/hour) dose
or 137Cs γ -rays
rate resulted in significant decreases in peripheral WBC and lym-
as the reference radiations. In male ICR mice irradiated with 2
phocyte counts as early as 4 hours post-irradiation
Gy of 70-MeV protons or 21-MeV electrons, peripheral WBC, lym-
At 24 hours post-irradiation with the 30.63–74.52 MeV pro-
phocyte and granulocyte counts decreased significantly in the ir-
tons, the dose response curves were nearly identical between the
radiated mice as compared to the sham irradiated controls, but
mice irradiated at the low and high dose rates for WBCs or lym-
the differences between the proton and electron irradiated mice
phocytes. In a separate experiment performed in the same series
were not statistically significant for WBCs, lymphocytes or granu-
of experiments, the mice were irradiated with protons at a very
locytes In 5 to 6-week old female ICR mice
low dose rate of 0.28 cGy/minute (17 cGy/hour), and the results
irradiated with 60Co γ -rays, 50-MeV protons or 70-MeV protons
were similar to those of the mice irradiated at the high dose rate
at low (0.5 Gy/hour) or high (0.5 Gy/minute) dose rate, the pe-
These results again indicate that dose rate
ripheral WBC count decreased in a dose-dependent manner at 24
in the range evaluated has little or no impact on the suppressive
hours post-irradiation and the RBE values for 50-MeV and 70-MeV
effects of proton radiation on the peripheral WBC or lymphocyte
protons at either low or high dose rate were not significantly
counts in irradiated mice.
different from 1 with 60Co γ -rays as the reference radiation at
During space missions, astronauts are potentially exposed to
the corresponding dose rates In a separate
SPE radiation in a reduced gravity environment. Thus, several ex-
study performed with 6-week old female ICR mice, the neutrophil
periments have been performed to determine the effects of sim-
count was monitored for 30 days post-irradiation with 0.5, 1 or
ulated microgravity on blood cell counts with or without addi-
2 Gy of 137Cs γ -rays or 74-MeV protons at dose rates of 0.44
tional exposure to space radiation [e.g., To
Gy/minute and 0.5 Gy/minute, respectively
A.R. Kennedy / Life Sciences in Space Research 1 (2014) 10–43
significantly at 48 hours post-irradiation (p < 0.001), both in adose-dependent manner.
In addition to the radiation dose, the dose rate also affected the
WBC and neutrophil counts at 3 hours, but not at 48 hours post-irradiation, when the WBC and neutrophil counts in the groupsirradiated at HDR were lower by approximately 16% and 32%, re-spectively, as compared to the WBC and neutrophil counts in thegroups irradiated at the LDR The dose rateeffects for the WBC and neutrophil counts were small, as com-pared to the magnitude of the effect of radiation dose at 3 hourspost-irradiation, and disappeared by 48 hours post-irradiation, sug-gesting that the higher dose rate might have only accelerated theonset of the radiation effects, but did not affect the overall mag-nitude of the radiation effects that developed at the later timepoints. Given the fact that the dose rate effect was not observedat 48 hours post-irradiation, when more pronounced losses of cir-culating WBCs, neutrophils and lymphocytes were observed in the
Fig. 2. Changes in neutrophil counts of mice exposed to a 2 Gy dose of SPE proton or
irradiated ferrets, the dose rate effect probably did not have a
γ -ray radiation. As shown in the figure, the effects of SPE like proton radiation on
biologically meaningful impact on the blood cell counts in the ir-
circulating neutrophil counts of mice are approximately the same as those of γ -ray
radiated animals.
radiation. In this figure, the absolute neutrophil counts are given at various times
RBE values were determined for 110 MeV protons using 60Co
post-irradiation. In radiation therapy patients, when the white blood cell countsfall below the level of 500 cells/microliter, it would trigger a medical response and
γ -rays as a reference radiation and peripheral leukocyte counts in
the patients would be considered as candidates for countermeasures (e.g., Neulasta
the irradiated ferrets as the biological endpoints. The RBE values
treatment). Thus, after irradiation of the mice with either SPE-like proton or γ -ray
derived from the WBC counts for 110 MeV protons delivered at the
radiation, the neutrophil counts fall to critically low values (< 500 cells per mi-
high or low dose rates at 3 and 48 hours post-irradiation ranged
croliter). (Courtesy of Dr. Ana Romero-Weaver; data from
from 1.19 to 4.02 at 0.75 Gy and declined with the increase in theradiation dose to a narrow range of 0.59 to 1.04 at 2 Gy
For the 2 Gy γ -ray and proton dose groups, the neutrophil
The RBE values calculated from the lymphocyte count
counts decreased by approximately 65% and 70%, respectively, at
data for the 110 MeV protons at the same time points and dose
the first nadir, which occurred on Day-4 post-irradiation, and then
rates were within a range of 0.83 to 1.41 at 0.75 Gy and showed a
fully recovered by Day-10 post-irradiation to levels that were not
slightly downward trend with the increase in the proton radiation
significantly different from the pre-irradiation control level. The
dose to a narrow range of 0.67 to 0.84 at 2 Gy. With only a few
neutrophil counts for the 2 Gy γ -ray and proton dose groups de-
exceptions, a similar downward trend was also observed for the
creased again by approximately 80% and 75%, respectively, at the
RBEs based on the neutrophil, monocyte and eosinophil counts in
second nadir, which occurred by Day-16 post irradiation, and then
the irradiated ferrets. These results suggest that 110-MeV protons
fully recovered again by Day 23 post-irradiation. Comparable re-
might be more effective than 60Co γ -rays in reducing the periph-
sults were observed in similar studies utilizing 74-MeV protons
eral leukocyte counts at the low end of the radiation dose range
and 137Cs γ -rays in which the changes in WBC counts were evalu-
evaluated in ferrets.
While the early radiation induced changes in the blood cells
ated over time The results from the
of ferrets were similar to those in the irradiated mice, the later
studies in mice indicate that proton radiation is not more effective
portion of the time course was quite different between mice and
than the commonly used reference radiations with respect to ef-
ferrets. In the irradiated mice, the initial sharp decrease in WBC
fects on hematopoietic cells
counts post-irradiation is followed by a gradual recovery to base-
Overall, the results show that the RBE values are not signifi-
line levels by 30 days post-irradiation. However, the recovery in
cantly different from 1 when the effects of SPE-like protons are
the WBC counts did not occur in ferrets exposed to radiation at
compared to those from the reference radiations (γ -ray or electron
doses of 1.5–2 Gy; 2 Gy was the highest dose of proton or gamma
radiation) in mice, and that SPE-like protons and γ -ray radiation
radiation evaluated in ferrets. This was shown to be due to the de-
result in almost identical dose–response curves over time, as illus-
velopment of disseminated intravascular coagulation
trated in [data in this figure are from
as discussed below.
2.1.3. Changes in peripheral leukocyte counts in Yucatan minipigs after
2.1.2. Changes in peripheral leukocyte counts in ferrets after irradiation
In ferrets irradiated with up to 2 Gy of 60Co γ -rays or 110-MeV
In the Yucatan minipigs irradiated with 6-MeV electrons at a
protons at a high dose rate (HDR) of 0.5 Gy/minute or a low dose
total skin dose of 25 Gy, the WBC count decreased significantly
rate (LDR) of 0.5 Gy/hour, the white blood cell (WBC) count de-
on day-1 post-irradiation, and then recovered by day-7 and in-
creased significantly within 3 hours after the radiation exposure,
creased significantly above the pre-irradiation level by day-30 after
and the average magnitude of the WBC decrease in the groups
irradiation The WBC count did not change
irradiated with 2 Gy of γ -ray or proton radiation at 48 hours post-
significantly in any other dose groups irradiated with electrons at
irradiation was approximately three times the decrease observed
a dose of 15 Gy or below. The lymphocyte count in the minip-
at 3 hours post-irradiation The lymphocyte
igs decreased significantly as early as 4 hours post-irradiation with
count also decreased significantly within 3 hours after irradiation
6-MeV electrons at skin doses of 15 or 25 Gy and then recov-
with 60Co γ -rays or 110-MeV protons, but the magnitude of the
ered to the pre-irradiation level by day-14 and day-7 for the 15-Gy
decrease was similar at the two time points (3 and 48 hours)
and 25-Gy dose groups, respectively. The neutrophil count did not
post-irradiation. In contrast, the neutrophil count increased signif-
change significantly after exposure to the electron irradiation in
icantly at 3 hours post-irradiation (p < 0.001) and then decreased
any dose groups at any time points except for a two-fold increase
A.R. Kennedy / Life Sciences in Space Research 1 (2014) 10–43
in the neutrophil count in the 25-Gy dose group at day-30 afterirradiation.
In the Yucatan minipigs exposed to radiation with 6 + 12 MeV
electrons, which is a suitable reference radiation with compara-ble body dose distribution as the SPE radiation the WBC count decreased significantly within a day post-irradiation in the 10, 15 and 20 Gy dose groups, but not in thegroups irradiated at a dose of 7.7 Gy or below Between day-1 and day-7 post-irradiation, the WBC countreached the lowest level, and then recovered slowly thereafter. Byday-30, the WBC counts in the 10, 15 and 20 Gy dose groups allrecovered to levels that were not significantly different from thebaseline value. The significant decrease in lymphocyte counts oc-curred earlier, to a greater extent and extended to lower dose (i.e.,5, 7.5 and 7.7 Gy) groups than the decrease in the WBC count afterthe 6 + 12 MeV electron irradiation. Within a day after irradiationwith 6 + 12 MeV electrons at doses up to 20 Gy, the lymphocytecount decreased by up to 77%. By day-30 post-irradiation, the lym-phocyte count in the 5, 7.5, 7.7 and 15 Gy dose groups recovered
Fig. 3. Changes in WBC counts of pigs exposed to eSPE or pSPE radiation with a skin
dose of 7.7 Gy. The pSPE dose involved several different energies of SPE radiation, as
to levels that were not significantly different from the baseline
described previously and the electron exposure involved two
value; however, the lymphocyte counts in the 10 and 20 Gy dose
different energies of electrons (6 and 12 MeV electrons to result in a dose distribu-
groups were still significantly below the baseline level. The neu-
tion like that expected for the SPE radiation exposure). The last time point for the
trophil count in the Yucatan minipigs irradiated with 6 + 12 MeV
evaluation of the changes in the circulating white blood cell numbers was 30 dayspost-irradiation. It can be observed in the figure that pig white blood cells return to
electrons did not show a consistent pattern of change among dif-
normal levels by 30 days post-exposure to electron radiation, but do not return to
ferent dose groups post-irradiation. Both increases and decreases
normal levels in proton irradiated pigs over the same time period. Significant dif-
in the neutrophil counts were observed at different time points
ference as determined by one-way ANOVA followed by the Tukey Test is indicated
post-irradiation with 6 + 12 MeV electrons at skin doses up to 15
by ∗ (p < 0.05), ∗∗ (p < 0.01) or ∗ ∗ ∗ (p < 0.001).
Gy, although the changes did not reach statistical significance dueto the relatively large variations in the control group and in some
to proton simulated SPE radiation do not return to normal levels
of the irradiated groups.
over this time period.
In the Yucatan minipigs irradiated with SPE-like protons with
Based on the WBC, lymphocyte and neutrophil count data for
energies of 155 MeV or below, the WBC count decreased signifi-
the minipigs exposed to 6 + 12 MeV electron radiation
cantly within a day post-irradiation with a single skin dose of up
and simulated proton SPE radiation
to 10 Gy Between day-1 and day-4 post-
RBE values were calculated for the effect of simulated pro-
irradiation, the WBC count reached the lowest level, and then
ton SPE radiation on leukocytes in irradiated animals. The results
recovered slowly thereafter. By day-30, the WBC count was no
show that the RBE value for the simulated SPE radiation varied
more than 38.3% below the baseline level for the animals irradi-
with both the radiation dose and the time post-irradiation
ated with 5, 7.7 or 10 Gy doses of protons. By day-90, the WBC
For WBC counts, the RBE calculated for the simu-
count recovered fully for the 5 Gy radiation dose group while re-
lated SPE radiation displayed a downward trend with the increase
maining 18.7% and 33.5% below the baseline level for the 7.7 and
in radiation dose on Day-1, Day-4 and Day-14 post-irradiation. At
10 Gy radiation dose groups. Significant decreases in lymphocyte
the 5 Gy proton dose level, the RBE values for the simulated SPE
counts occurred earlier and to a greater extent than the decrease
radiation were 2.0, 4.1 and 3.3 on Day-1, Day-4 and Day-14, re-
in the WBC count for the minipigs irradiated with 5, 7.7 and 10
spectively, after irradiation For lymphocyte
Gy of protons. On day-1 after irradiation, the lymphocyte count
counts at 4 hours post-irradiation, the RBE for the simulated SPE
reached the lowest level, which was 73.0%, 79.7% and 89.5% below
radiation also showed a downward trend with increasing dose,
the baseline level for the 5, 7.7 and 10 Gy radiation dose groups,
with the RBE value changing from 9.6 at 5 Gy to 4.6 at 10 Gy.
respectively. By day-30 post-irradiation, the lymphocyte count in
However, the RBE trend for the simulated SPE radiation calculated
the irradiated animals was not more than 34.5% below the base-
from the lymphocyte count data was relatively flat on Day-1 and
line level. By day-90, the lymphocyte count recovered fully for the
Day-4 and only slightly downward with the increase in the radia-
5 Gy radiation dose group and was not more than 19.5% below the
tion dose on Day-14 post-irradiation. For neutrophils, the RBE for
baseline level for the two higher radiation dose groups.
the simulated SPE radiation also displayed a noticeable downward
The neutrophil count change in the Yucatan minipigs displayed
trend with the increase in radiation dose on Day-4 and Day-14.
quite a different time course as compared to the changes ob-
The fitted RBE values were higher than 1.00 at all three simulated
served in the WBC and lymphocyte counts. The neutrophil count
SPE radiation dose levels of 5, 7.7 and 10 Gy for WBCs and lym-
increased by up to 79.8% at 4 hours post-irradiation, and then de-
phocytes, and the lower limits of the 95% confidence interval for
creased by up to 42.1% on day-1 after irradiation. The neutrophil
the RBEs were above 1.00 for all dose levels on Day-1, 4 and 14
count reached the lowest level between day-4 and day-14 post-
except for 10 Gy on Day-1 for WBCs and 7.7 and 10 Gy on Day-4
irradiation, and then recovered slowly thereafter. By day-90 post-
for neutrophils. In addition, the ED10 and ED50 values for the sim-
irradiation, the neutrophil counts in the 7.7 Gy and 10 Gy dose
ulated SPE proton radiation were significantly lower than those for
groups were still 29.3% and 48.0% below the pre-irradiation level.
the 6 + 12 MeV electron radiation on Day-1, 4 and/or 14 post-
The results for the pig WBC counts over a 30 day experimental pe-
irradiation for WBCs, lymphocytes and/or neutrophils
riod following exposure to proton or electron radiation are shown
These results indicate that simulated SPE proton radia-
in It should be noted that, following exposure to electron
tion is significantly more effective than 6 + 12 MeV electrons with
simulated SPE radiation, the WBC counts return to normal levels
respect to the effects on the peripheral WBCs, lymphocytes and
by 30 days post-irradiation, but the WBC counts in pigs exposed
neutrophils, especially at the low end of the radiation dose range
A.R. Kennedy / Life Sciences in Space Research 1 (2014) 10–43
evaluated. It was observed that the neutrophils ware particularly
radiation may be far more hazardous to humans than previously
sensitive to the damaging effects of proton radiation.
estimated from studies performed with small animals.
2.1.4. Summary of effects of SPE radiation on hematopoietic blood cell
2.2. Immune system effects
counts in ferrets, mice and pigs
For hematopoietic cells, significant decreases in white blood
2.2.1. Effect of SPE-like radiation on gastrointestinal tract integrity
cell counts were observed in mice and ferrets irradiated at high
Numerous studies have been performed to evaluate the effects
(∼0.5 Gy/minute) and low (0.5 to 0.17 Gy/hour) dose rates, starting
of SPE radiation on the immune system as a part of research on the
at doses of 0.5 Gy and up to 2 Gy. In some, but not all, of the stud-
acute risks of SPE radiation exposure
ies involving dose-rate comparisons, lowering the dose rate had a
small, but statistically significant, sparing effect on the white blood
the SPE radiation studies were performed with and without mi-
cell count parameters evaluated, but in many other dose-rate ex-
crogravity simulated with PWS or HS
periments performed, such changes were not observed. Thus, over
the range of dose-rates evaluated, it is concluded that lowering the
For the studies related to immune sys-tem effects, mice were exposed to homogeneous doses of either
radiation dose-rate does not produce sparing effects on hematopoi-
γ -ray or SPE-like proton radiation. The effects of γ -ray and SPE-
etic cell counts that are of biological significance. At the higher
like proton radiation were comparable in these studies, and none
doses of proton or γ -ray radiation, the neutrophil counts in the
of the observed effects described below were specific to proton or
blood of both mice and ferrets reach critically low levels (< 500
γ -ray radiation.
cells per microliter)
The gastrointestinal (GI) tract contains over 1012 bacteria, and
If such a low value occurred in a patient in a hospital
these bacteria have many important functions including carbohy-
(e.g., following radiation or chemotherapy for cancer), this would
drate fermentation and absorption, repression of pathogenic mi-
trigger a medical response, and suggest the use of countermea-
crobial growth, and continuous modulation of the gut and sys-
sures to increase the level of neutrophils. For the pigs, there were
temic immune system. A critical function of the GI tract is the
also highly significant reductions in the levels of WBCs following
containment of commensal bacteria, which involves the control
proton or electron SPE radiation exposure, but they did not reach
of bacteria and bacterial product passage across the GI mucosa,
the critically low values observed in the mice or ferrets at any of
known as bacterial translocation; this function can be disturbed in
the high skin doses evaluated.
many different diseases. In a study performed with ICR mice at
A major difference in the pig response to the electron and pro-
5–6 weeks of age, irradiation with 2 Gy of 50 or 70-MeV protons
ton SPE radiation was that the neutrophil count did not show a
resulted in a transient increase of lipopolysaccharide (LPS) in the
meaningful recovery by 3 months after exposure to 10 Gy of pro-
serum at one day post-irradiation, and the increase was accompa-
ton SPE radiation . For pigs exposed to 10 or
nied by increases in acute-phase reactants, such as lipopolysaccha-
20 Gy of electron SPE radiation, the neutrophil count recovered as
ride binding protein (LBP) and soluble CD14 (sCD14), circulatory
expected within a month after the radiation exposure
pro-inflammatory cytokines (including TNF-α, IL-1β and IL-6), and
These results indicate that the pigs might be less ca-
transient disruption of tight junctions in the GI track, which in-
pable of repairing the DNA damage caused by the proton radiation
dicated a transient increase in bacterial translocation across the
exposure than the DNA damage caused by the electron radiation at
GI tract and systemic activation of the innate immune system
similar or higher doses. Similar results were observed for mice ex-
HS was also shown to cause a breakdown in the
posed to proton total body radiation at a dose of 5.5 Gy, which
containment of Gram negative bacterial products, as measured by
significantly suppressed the neutrophil count at 9 weeks post-
circulating LPS The combined treatment of a 2 Gy
irradiation In contrast, the neutrophil count
dose of either γ -ray or SPE-proton radiation with HS led to a
in mice exposed to x-rays at comparable or considerably higher
greater and more sustained elevation in the level of LPS in the
doses (up to 8 Gy) was fully recovered at 8 weeks post-irradiation
serum. Bacterial translocation is known to increase circulating lev-
These results suggest that mouse neutrophils
els of LPS and other bacterial components, which include bacterial
might also be more sensitive to the DNA damaging effects of pro-
DNA To determine whether the increase in
ton radiation when given at a relatively high dose (5.5 Gy). At a
LPS induced a systemic response, LBP, which is a type 1 acute
dose of 2 Gy, the neutrophils in the irradiated mice appeared to
phase protein, was measured in ICR mice exposed to radiation
be equally sensitive to the damage from proton and γ -ray irradia-
with and without hindlimb suspension. LBP is a circulating pro-
tein that binds to LPS of Gram-negative bacteria; it is constitutively
We have calculated RBE values for SPE-like radiations using
present but can be induced to higher levels during various types
hematopoietic cell count data from mice, ferrets and Yucatan
of infection and inflammatory processes. LBP was increased after
minipigs irradiated with SPE-like proton radiation and a suitable
treatment with proton radiation or HS and was increased further
reference radiation (e.g., γ -rays, x-rays or electrons). A higher RBE
when these stressors were combined Similar
value for a given biological endpoint, such as blood cell count, indi-
results were observed for sCD14, another very sensitive marker of
cates that the SPE-like proton radiation is more effective than the
increased levels of circulating LPS. Circulating levels of interferon-
reference radiation in affecting that biological endpoint. In these
alpha (IFN-α) were measured, and at least additive levels of IFN-α
studies, it was observed that there were: 1) different RBEs for dif-
were observed for mice treated with both radiation (2 Gy of γ -ray
ferent biological endpoints in the same animal species/strain, and
or proton radiation) and HS. These results demonstrate that cir-
2) different RBEs for the same endpoint in different species/strains.
culating LPS, resulting from exposure to SPE-like radiation, HS or
The RBE values estimated based on the WBC results vary greatly
both, led to a systemic response. It has been concluded from these
between mice, ferrets and pigs, with the RBE values being greater
studies that there is a synergistic effect when hindlimb-suspended
in ferrets than in mice at times up to 48 hours post-irradiation
mice are additionally exposed to SPE-like radiation.
and considerably greater in pigs than in
To determine the mechanisms involved in the increased bacte-
ferrets and mice This trend suggests that
rial translocation across the GI tract, immunohistochemical stain-
the RBE values for WBC counts in humans could be considerably
ing for the tight junction protein, Claudin-3, was performed on
greater than those observed in smaller mammals, and SPE proton
terminal ileum sections of mice, and a significant increase in the
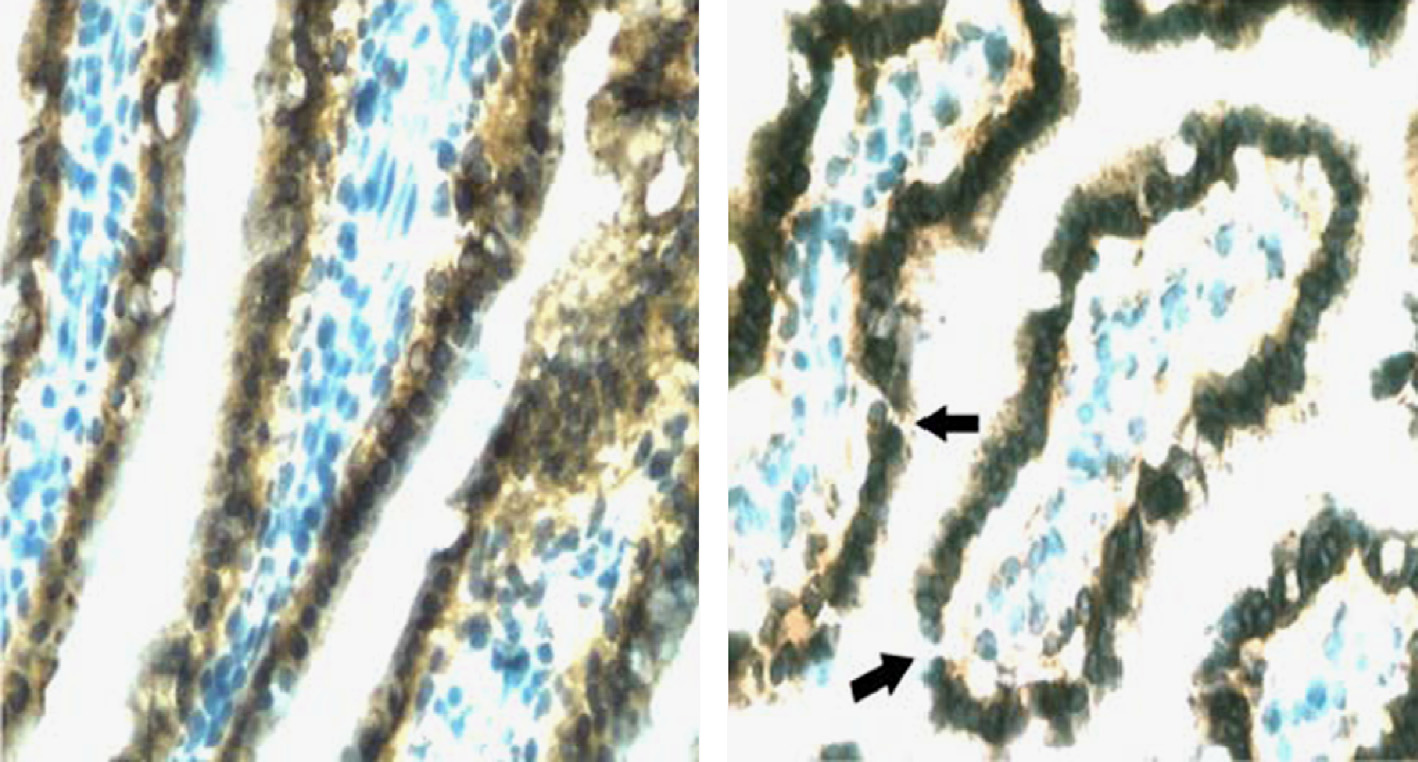
A.R. Kennedy / Life Sciences in Space Research 1 (2014) 10–43
Fig. 4. Proton radiation induces breaks in the GI epithelial barrier. Terminal ileum obtained 2 days post-irradiation from a control mouse (A) or a mouse irradiated with
2 Gy of protons at the low dose rate (B), and stained for Claudin-3. Black arrows show regions of tight junction incongruity. Original magnification 400×. Reprinted with
permission from Radiation Research
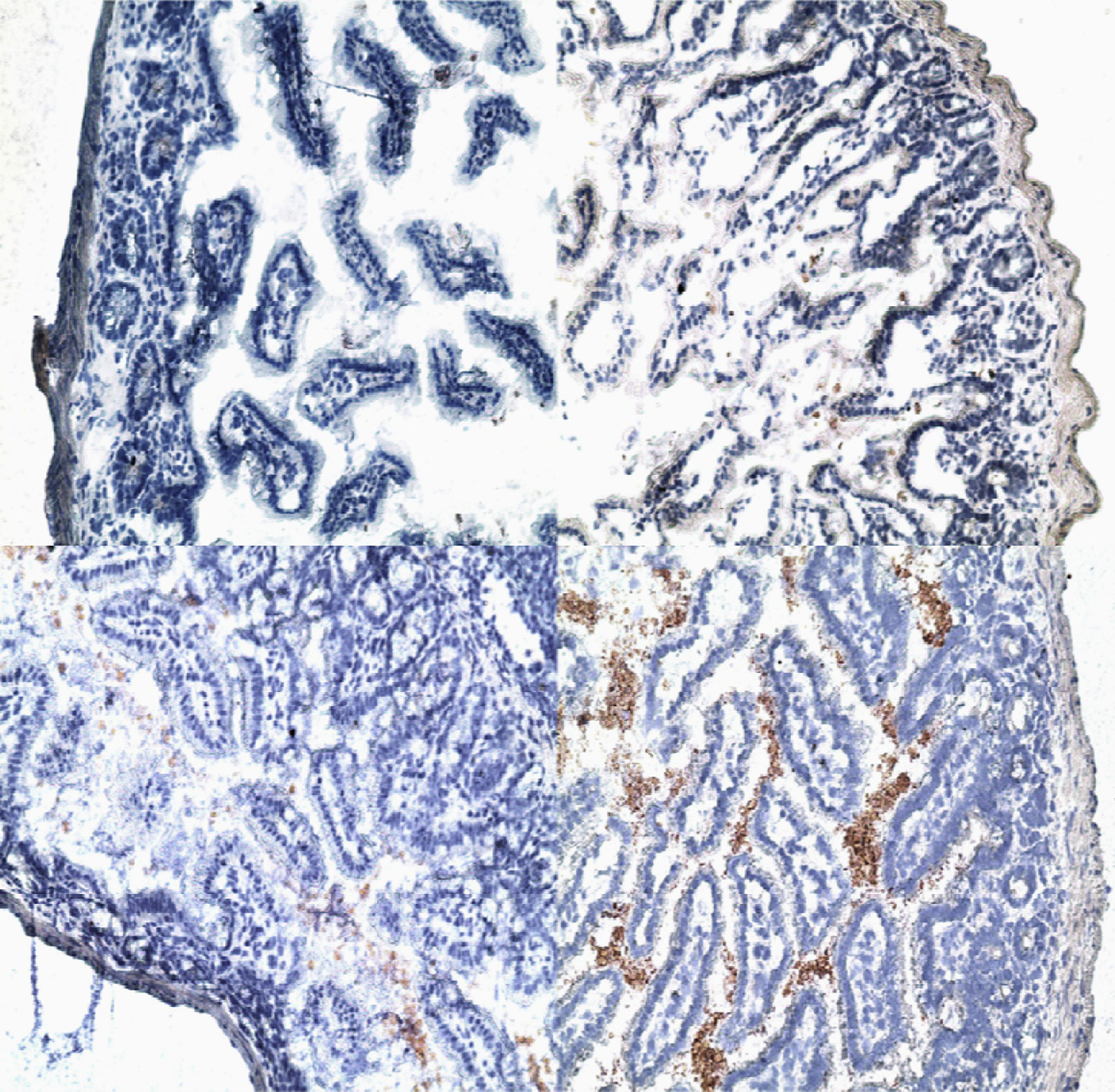
Fig. 5. SPE-like proton radiation and hindlimb suspension lead to the accumulation of LPS in subepithelial regions of the ileum. Terminal ileum obtained 4 days post irradiation
and/or 6 days post hindlimb suspension or from control animals was stained for LPS using a mouse mAb specific for E. coli LPS. A: represents ileum from a control mouse,
B: ileum for a mouse irradiated with 2 Gy of 70 MeV protons, C: ileum from a mouse subjected to hindlimb suspension, and D: ileum from a mouse subjected to HS and
irradiated with 2 Gy of 70 MeV protons. It can be observed in the figure that the amount of LPS accumulated in the subepithelial region of the ileum is considerably greater
in the mouse exposed to both HS and SPE proton irradiation than in mice exposed to either HS or SPE proton radiation alone. Original magnifications – 200×.
A.R. Kennedy / Life Sciences in Space Research 1 (2014) 10–43
number of breaks and reductions in staining were observed in
erance", in which subsequent responses are reduced in quality and
mice exposed to proton or γ -ray radiation and HS, as illustrated in
quantity, which is thought to protect the host by limiting excessive
These studies indicate that SPE-like radiation and hindlimb
inflammation and preventing septic shock
suspension induced breaks in the GI epithelial barrier, and sug-
As has been observed in many different dis-
gest that the increased frequency of breaks could be responsible
mechanistically for the increase in translocation of bacterial prod-
ucts. To further substantiate this association, the terminal ileum
continuous or extended exposure to PAMPs
was stained with two antibodies that recognize LPS, a mouse mon-
can lead to long-lived immune dysfunction. This condition may ex-
oclonal antibody against E. Coli (j5) LPS and a goat anti-lipid A
ist during space flight, as LBP, a well-known marker of immune ac-
IgG that cross-reacts with Pseudomonas aeruginosa, Klebsiella pneu-
tivation, is known to be elevated in astronauts' plasma
moniae, E. coli 0157, Salmonella enteriditis, Enterobacter aerogenes,
It is possible that the elevated blood
E. Hermanii, Yersinia enterocolitica and Shigella sonnei. Control an-
levels of LBP resulting from exposure to SPE radiation and simu-
imals demonstrated no LPS specific staining in the intervillous
lated microgravity (HS) may bring about immune dysfunctions of
space, while low levels were observed in mice treated with SPE-
the sort that have been observed during and after extended space-
proton radiation or HS alone. For the mice exposed to the com-
bined treatment with SPE-proton radiation and HS, there was ahigh level of diffuse staining, as shown in Thus, the conclu-
2.2.2. Effect of SPE-like radiation on skin immune function
sion from these studies was that SPE-like radiation and hindlimb
Two models were used to evaluate the effects of SPE radiation
suspension induced the accumulation of LPS in subepithelial re-
on skin immune function. One involved Yucatan minipigs, as their
gions of the ileum.
skin histological structure is an accurate model of human skin, and
It has been estimated that astronauts could receive a dose of up
the other involved mice, in which in depth investigations of alter-
to 2 Gy to the bone marrow from SPE radiation
ations in immune function could be performed. A major difference
As discussed above, when a 2 Gy dose of radi-
between the models used involves the depth of radiation penetra-
ation is combined with simulated microgravity, an enhanced and
tion. The pigs were exposed to electrons or protons as an SPE-like
prolonged impairment of commensal bacteria containment was
inhomogeneous dose of radiation, with a relatively high dose de-
observed. We have identified the mechanism for the loss of con-
livered to the epidermis and dermis and lower doses delivered to
tainment, which is that radiation plus HS leads to breaks in the
internal organs while the mice received proton or
tight junctions between GI tract epithelial cells, which results in
reference radiation (γ -rays) exposures as a homogeneous dose of
the migration of LPS into the subepithelial tissue. Potential ther-
radiation. The pigs were exposed to skin doses as low as 2.5 Gy to
apies to treat this immune defect could target the GI defect that
up to 25 Gy, which was the highest skin dose evaluated in these
leads to bacterial translocation or by reducing the inflammatory
studies. It should be pointed out that skin doses for major histori-
activity of translocated bacterial products. The mucosal integrity of
cal SPEs were calculated to be as high as 32 Gy
the GI tract is maintained by a population of CD4+ cells that pro-
however, such a high skin dose would represent a worst case sce-
duce IL-17 (Th17 cells). Their loss is known to be correlated with
nario that would not be expected to occur frequently. Delayed type
increased bacterial translocation Although
hypersensitivity (DTH) responses to phytohemagglutinin (PHA) and
there are no current therapies that can mitigate the loss of GI Th17
LPS were measured prior to radiation and at 7, 14 and 30 days
cells, this is an area of research worthy of investigation. Antibiotics
post-irradiation. The responses were symmetric and recorded as
can be used to treat the increased bacterial translocation, and they
the average distance across for induration, erythema and ulcera-
are known to be capable of reducing serum LPS levels
tion. Since a similar pattern was observed for all doses of radiation
used, with significant increases in the response after radiation, but
Numerous immune system alterations have been associated
with no dose dependency, we analyzed all radiation dose groups
with space flight in humans and in animals during ground-based
together for the response to control (PBS), PHA and LPS treatments.
spaceflight models (e.g. HS), as has been reviewed [e.g.,
A significant enhancement in the DTH response to PHA was ob-
served at all post-irradiation time points evaluated. The responses
The major effects of spaceflight on the immune system
to LPS were not significantly elevated at day 7, but they become
have been well-characterized, and include changes in cytokine
statistically significant at 14 and 30 days post-irradiation
production, leukocyte subset distribution and antibody production
The appearance of ulceration after radiation expo-
Examples of cytokines released
sure was noted for both PHA and LPS treatments. It is assumed
in response to stimulation include the following: an increase in
that ulceration occurred as part of the enhanced immune response
anti-inflammatory cytokines and a decrease in TNF-a in LPS stim-
post-irradiation. If radiation was responsible for the ulceration, ul-
ulated spleen cells reductions in interferon-γ
ceration should have increased with increasing doses of radiation,
and IL-2 following phorbol 12-myristate 13-acetate and ionomycin
which was not the case. Mice were exposed to a homogeneous
stimulation of peripheral blood cells of astronauts
dose of radiation up to a 2 Gy dose. Mouse skin challenged with
and reduced NK cell number and function
intradermal PHA was measured and a similar increase in DTH re-
Such alterations in im-
activity was noted after exposure to 2 Gy of proton or γ -ray radi-
mune function are similar to those brought about when there is
ation and (Weissman, D., Unpublished data)].
increased immune activation produced by exposure to pathogen
Skin is known to contain high frequencies of FoxP3 expressing
associated molecular patterns (PAMPs), such as LPS
regulatory CD4+ T cells (Tregs). In the skin from irradiated pigs
and mice, immunohistochemical analysis demonstrated a loss of
CD3+ and CD25+ cells. To determine whether loss of Tregs was re-
cific examples of this are as follows: a reduction in proinflamma-
sponsible for the enhanced DTH responses, RNA was isolated from
tory cytokine production by myeloid cells
murine skin prior to and at 2, 7 and 14 days post-irradiation for
a reduction in antigen-specific T cell effector
quantitative PCR measurements of CD3, FoxP3 and GAPDH mRNA.
cytokine responses and a reduc-
The results demonstrate that there was a statistically significant
tion in circulating NK cells The changes in
reduction in FoxP3 mRNA at all 3 time points following expo-
responses brought about by exposure to PAMPs are known as "tol-
sure to 2 Gy of irradiation. Smaller decreases in CD3+ cells were
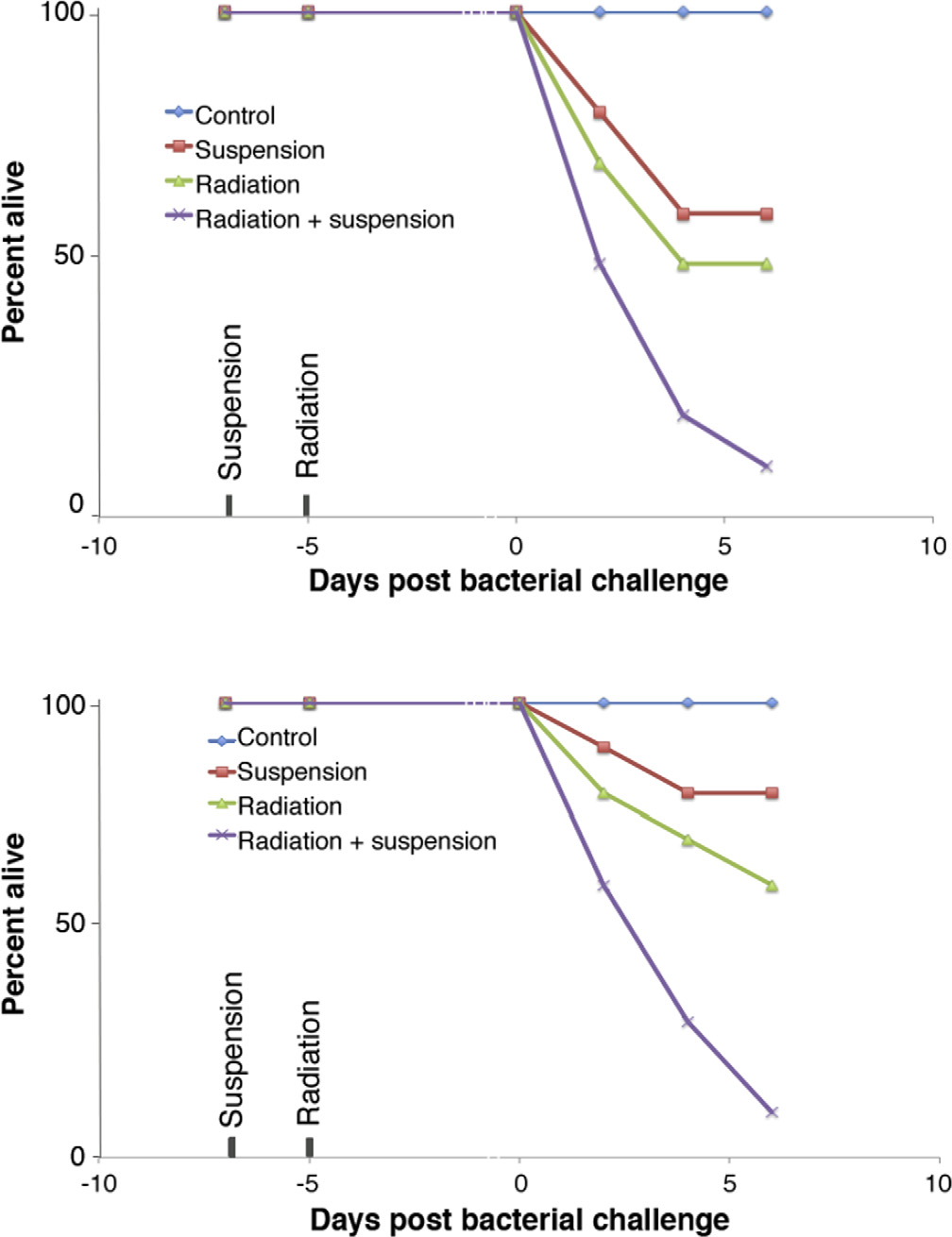
A.R. Kennedy / Life Sciences in Space Research 1 (2014) 10–43
observed, demonstrating that FoxP3 positive cells were being se-lectively lost (Zhou, Y., Ni, H., Balint, K., Sanzari, J.K., Dentchev, E.,Diffenderfer, E., Wilson, J., Kennedy, A.R., Cengel, K.A. and Weissman, D.,Unpublished data). Mouse skin was obtained at various time pointspost-irradiation in the experiment and single cell suspensions oflymphoid cells were obtained by enzymatic digestion for analysisof CD4, CD25 and FoxP3 cells by flow cytometry. Statistically signif-icant decreases in the percent of CD4+ T cells expressing CD25 andFoxP3 were observed at all post-irradiation time points evaluated.
The greatest loss was observed at 4 days post-irradiation, with aslow increase in Tregs over the next 28 days. The proliferation ofskin CD4+ T cells increased with the loss of Tregs, demonstratinga functional effect.
The loss of skin Tregs could be due to the cell killing effects
of radiation or alterations in trafficking. We examined splenic lym-phoid cells and observed that, with the loss of skin Tregs post-irradiation, there was a statistically significant increase in the per-cent of Tregs in the spleen. The increase in Tregs led to a drop inthe proliferation of activated CD4+ and CD8+ cells. A recent reporthas indicated that Tregs traffic through skin and that they are themain types of cells exiting skin during inflammation In these studies, it was observed that half of the skin cellsthat migrated to draining lymph nodes were Tregs at steady state.
Tomura et al. also noted that when an immune reaction was in-duced in the skin, the frequency of Tregs draining to lymph nodesincreased significantly and made up the majority of cells exitingthe skin In addition, it was found that theincrease in Tregs leaving the skin resulted in more suppression ofT cell activation in the draining lymph nodes and spleen whereTregs accumulated.
To identify mechanisms for the radiation induced depletion of
skin T cells, inflammatory genes in mouse skin obtained prior toand at various time points after mice were treated with a sin-gle radiation dose of 2 Gy were analyzed. It was observed that
Fig. 6. Survival of mice challenged with bacteria after irradiation with or without
at 6 hours post-irradiation, multiple acute inflammatory markers,
hindlimb suspension. C3H/HeN mice were treated with HS, 2 Gy of SPE-like proton
CXCl chemokines, were upregulated. At 24 hours and continuing
radiation, or both, and then the mice were challenged with a non-toxic dose ofPseudomonas aeruginosa (Panel A) or Klebsiella pneumoniae (Panel B) bacteria. There
through day 14, chronic inflammatory markers, CCL chemokines,
was a high level of mortality observed in all treatment groups except the control
complement and IL-10, were induced. These data indicated that
group; these mice were not exposed to HS or radiation, but were given the bacterial
irradiation at a dose of 2 Gy induces long-lived inflammatory
challenge. (The images were provided by Drew Weissman, M.D., Ph.D. [data from
changes in the skin, including alterations in chemokines known to
attract Tregs to the site of infection, including CCL17 and CCL22which are down-regulated. We hypothesize
fection in an astronaut during the Apollo 13 mission
that the radiation induced inflammation establishes an environ-
ment that induces T cells, namely of the regulatory phenotype, to
As the control of infections during spaceflight is
leave the skin and reduces their ability to return to the skin. The
a major problem, much effort has been focused on determining
lack of Tregs in the skin likely results in a loss of control of the
the effects of spaceflight stressors, such as simulated microgravity
inflammatory response induced by PHA and LPS challenges, result-
and SPE-like radiation, on the ability to defend against a bacte-
ing in enhanced responses post-irradiation. The clinical significance
rial challenge In these studies, mice were ex-
of this for an astronaut exposed to SPE radiation is unknown. Po-
posed to SPE-like radiation and/or HS, and then challenged with
tentially, as skin abrasions occur during space flight, an enhanced
Pseudomonas aeruginosa systemically or Klebsiella pneumoniae by
inflammatory response in the setting of reduced immune compe-
inhalation. Numbers of bacteria that allow most to all of the un-
tence due to Treg migration to lymphoid organs could result in a
treated animals to survive were used in these studies so that any
reduced ability to control an infection.
decrement in immune function could be measured by increasedamounts of bacteria in the blood and lung and morbidity. Three
2.2.3. Effect of SPE radiation and hindlimb suspension on immune
different strains of mice were used: ICR mice are an outbred strain
function measured by bacterial challenge
initiated in 1948 from Swiss mice, C3H/HeN mice are inbred and
Infections of the skin, eyes and respiratory tract are com-
have no known defects of polymorphisms that impair DNA re-
mon in astronauts: infections have been reported 13 times in the
pair or the response to ionizing radiation, whereas Balb/c mice
Apollo and 8 times in Skylab missions, and spacecrafts need to be
have 2 different polymorphisms in DNA-dependent protein kinases
equipped with numerous antibiotics for treatment of such infec-
(DNA-PKcs) that mediate non-homologous end joining, which re-
tions A number of bacterial
sults in decreased, but not absent, function
infections have been observed in astronauts during or soon after
To measure the effect
missions, with organisms that do not typically lead to such infec-
of the HS stress and SPE-like radiation on the ability of differ-
tions in healthy people. As one example, Pseudomonas aeruginosa,
ent strains of mice to effectively clear a challenge with bacteria,
which does not ordinarily infect healthy people, was identified as
hindlimb suspended and/or irradiated mice were exposed 5 days
the pathogen that caused a serious life threatening urinary tract in-
later to Pseudomonas aeruginosa or Klebsiella pneumoniae. The mice
A.R. Kennedy / Life Sciences in Space Research 1 (2014) 10–43
were followed daily for signs of systemic and pulmonary infec-
likely to be less effective with repeated applications over long pe-
tions. The results for all three strains of mice were comparable
riods of time. Numerous infections have already been documented
and indicated that the mice exposed to HS and SPE radiation failed
in astronauts and are considered a major hazard for spaceflight;
to control a challenge with Pseudomonas aeruginosa or Klebsiella
some infections have been minor while others have been serious
pneumoniae, which led to a high morbidity/mortality rate
and life-threatening, including a debilitating dental infection and
as illustrated in [data from (note: in
an incapacitating urinary tract infection
these studies, morbidity was the same as mortality). Either SPE
radiation or HS alone had some effects on morbidity in these
It is known that spaceflight conditions alter the gene ex-
studies, but when combined, they led to almost complete mor-
pression patterns, virulence and virulence phenotypes of bacterial
bidity/mortality. Other studies in this series of experiments led
pathogens with evidence of increased vir-
to the following conclusions: 1) similar levels of morbidity were
ulence under space flight conditions; thus, longer space flights are
observed after the challenge with Pseudomonas bacteria in male
likely to lead to considerably more serious immunological prob-
and female mice, indicating a lack of gender/sex differences in
lems than observed so far in the space program.
this effect, 2) a dose of 1.5 Gy of total body radiation impaired
The other immunological issue of great importance to the space
the ability of mice to control the bacterial challenge in a fash-
program is the observation of additive or synergistic-adverse ef-
ion similar to that observed for 2 Gy with little difference in the
fects caused by SPE radiation exposure and simulated microgravity
morbidity observed between these two dose groups, but the mor-
conditions; interactive effects between SPE radiation and simulated
bidity differences observed for the 1 Gy dose group compared to
microgravity have been observed for various immunological pa-
those from the control group were not statistically significant, in-
rameters at doses of radiation that could be received by astronauts
dicating that the threshold dose for morbidity resulting from the
during space travel [e.g.,
bacterial challenge is between 1 and 1.5 Gy, 3) the relative in-
The results indicate that, under simulated
creases in morbidity were similar for all 3 strains of mice, sug-
microgravity conditions, the effects of a given dose of SPE radi-
gesting that the polymorphisms in DNA-PKcs in Balb/c mice did
ation can be considerably more severe than the effects observed
not significantly affect the response to a bacterial challenge, and 4)
for the same dose of radiation in normal, control animals. These
peripheral blood granulocyte counts were determined in C3H/HeN
results suggest the possibility that, in the space microgravity en-
mice challenged with Pseudomonas aeruginosa at time points prior
vironment, the effects of a given dose of SPE radiation could be
to HS and irradiation and before and after the bacterial challenge.
comparable to those observed for a significantly higher dose of ra-
In these studies, there was a reduction in the peripheral blood
granulocyte counts observed post-irradiation, which was similar tothat described previously
2.3. Emesis
The numbers of peripheralgranulocytes increased as expected after bacterial challenge in con-
The early phase of the acute radiation syndrome, which is
trol or irradiated mice, but the mice treated with HS, with or
known as the prodromal syndrome, can include nausea, retch-
without radiation, failed to elevate the blood granulocyte counts as
ing, vomiting, diarrhea, and fatigue
expected Similar blunting of the peripheral blood
These effects often manifest within 1 to 72 hours post-
granulocyte counts in response to systemic infection in HS animals
irradiation at sub-lethal doses, with a latency time inversely corre-
was also observed in Balb/c and ICR mice (Drew Weissman and
lated with dose. Vomiting is the reflexive act of forcefully ejecting
colleagues, unpublished data).
the stomach contents through the mouth by coordinated mus-cle contraction. Published clinical studies have demonstrated that
2.2.4. Summary of the effects of SPE radiation on the immune system
patients receiving total body irradiation or upper-abdominal irra-
The effects of SPE radiation on immune system parameters
diation often show nausea, retching and vomiting as side effects
are of great importance to the space program as they are poten-
tially life-threatening at doses that could conceivably be received
lation between retching and vomiting events has been established
by astronauts during space travel. It has been observed that ex-
in the ferret model Emetic
posure to SPE radiation along with HS, with additional exposure
responses to various pharmacological agents, cytotoxins and radi-
to a bacterial challenge, leads to a very high level of morbidity,
ation have been compared previously among humans and various
which is equal to mortality, in the studies performed in mice.
animal species including nonhuman primates, dogs, cats, and fer-
The bacterial challenge utilized bacteria known to be associated
rets Ferrets are considered to be a useful species in
with astronaut infections and are already part of the spacecraft
emesis research especially for radiation and
environment (Pseudomonas aeruginosa) or that are part of the nor-
cytotoxic drug-induced emesis and data
mal bacterial flora of the mouth, skin and intestines (Klebsiella
from the ferrets have been used by the Department of Defense to
pneumoniae); the bacterial challenges utilized in the mouse stud-
develop a mathematical model for the human emetic response to
ies described above were performed with bacterial levels that are
radiation Another advantage of the ferret
non-toxic (or minimally toxic) to the normal control mice. Un-
model is that the prodromal response appears at lower radiation
der the conditions described above leading to morbidity/mortality
doses and with an earlier onset time as compared to other species,
in the mice, death could be prevented by treatment with an an-
including humans Thus, we have chosen ferrets as an
tibiotic (enrofloxacin). Enrofloxacin is approved by the FDA as a
experimental model system to determine the effectiveness of pro-
veterinary antibiotic (marketed by the Bayer Corporation under the
tons at the energy, doses and dose rate ranges relevant to the SPE
trade name Baytril). A similar antibiotic (ciprofloxacin) is in wide
radiation exposures expected during space travel.
use in human populations. While the effectiveness of the antibi-
In our studies performed with female descented Fitch ferrets
otic is outstanding, the major problem associated with the use of
aged 12 to 16 weeks, irradiation with 60Co γ -rays or 155-MeV
antibiotics for bacterial infections in astronauts is that organisms
protons at a high dose rate of 0.5 Gy/minute resulted in dose-
resistant to the treatment can grow out in a very short period of
dependent changes in the endpoints related to retching and vom-
time [as short as a one-week period of time (Personal Commu-
iting, such as the fraction of animals that retched or vomited, the
nication, Drew Weissman, M.D., Ph.D.)]. Thus, it is expected that
number of retching and vomiting events, the length of the latency
in the exploration class missions of the future, the antibiotics are
period leading to the first retching or vomiting event and the dura-
A.R. Kennedy / Life Sciences in Space Research 1 (2014) 10–43
tion between the first and last retching or vomiting events
lial cells were exposed to 20 Gy irradiation with a 6-MeV elec-
A dose–response relationship was observed for ferret
tron beam at a fixed dose of 2.4 Gy/minute
vomiting and retching at the high dose rate. The minimum radia-
It has also been shown that in human peripheral blood
tion doses required to induce statistically significant changes in the
mononuclear cells (PBMNCs) after irradiation with a 6-MeV elec-
retching- and vomiting-related endpoints were 0.75 and 1.0 Gy, re-
tron beam, TF was up-regulated, which led to a significant in-
spectively; thus, these values are considered the threshold doses
crease in PBMNC-associated pro-coagulant activity over a time
for radiation induced retching and vomiting in the ferret model.
period of 7 days post-irradiation. Increased cellular TF protein
The RBE of the proton radiation at the high dose rate did not dif-
concentration was observed up to 7 days post-irradiation, and
fer significantly from 1. Similar, but smaller and less consistent,
microparticle-associated TF activity was increased significantly 3
changes in the retching- and vomiting-related endpoints were also
days post-irradiation as compared with the non-irradiated con-
observed for ferrets irradiated with γ -rays and protons delivered
trols. PBMNC-derived microparticles post-irradiation also initiated
at the low dose rate of 0.5 Gy/hour. Since this low dose rate is
the plasma clotting faster than microparticles derived from con-
similar to a radiation dose rate expected during an SPE, these re-
trols. The radiation induced TF expression and increase in proco-
sults suggest that the risk of SPE radiation-induced vomiting is low
agulability of PBMNCs and cell-derived microparticles may repre-
and may reach statistical significance only when the radiation dose
sent a possible mechanism by which ionizing radiation enhances
reaches 1 Gy or higher.
blood thrombogenicity . In a study per-
Ferrets have also been used previously to study emesis induced
formed with leukoreduced fresh-frozen plasma irradiated with
by radiation with 60Co γ -rays
30 Gy of γ -rays, prothrombin time (PT), activated partial throm-
0.6-GeV/n 56Fe ions, neutrons and 200-MeV
boplastin time (aPTT), thrombin time, antithrombin III, protein C,
protons and the emetic re-
protein S, vWF, ristocetin cofactor, plasminogen–α2-antiplasmin,
sponse in ferrets was found to be dependent on the type and dose
the coagulation factors fibrinogen, FII, FV, FVII, VIII, F IX, FX,
of radiation. High LET 56Fe particles and fission neutrons were
FXI, FXII, FXIII, and activated factor XII (FXIIa), D-dimer, fibrin
comparable in their ability to produce emetic responses (retching
monomer, thrombin–antithrombin complex, prothrombin fragment
or vomiting) in ferrets with an ED50 of 0.35 Gy and 0.40 Gy, re-
1 + 2 (F1+2), plasmin–α2-antiplasmin complexes, and platelet
spectively whereas γ -rays were shown to be
factor 4 were determined, and PT, aPTT and FVIII activities were
intermediately effective with an ED50 of 0.77 Gy to
found to be decreased significantly, whereas activities of the co-
0.95 Gy and high energy electrons were the
agulation factors FII, FV, FVII, FIX, FX, FXII were increased signifi-
least effective, with an ED50 of 1.38 Gy The
cantly post-irradiation
ED10, ED50 and ED90 values estimated for the fraction of animals
To determine whether SPE-like proton radiation could affect
that vomited after proton irradiation at the HDR were comparable
blood clotting times, we have performed experiments with 12
to the ED10, ED50 and ED90 values after γ -ray irradiation at the
to 16 week old descented female Fitch ferrets and demonstrated
high dose rate in our study or at a dose-rate
significant increases in the PT at 3 hours post-irradiation with
of 1 Gy/minute, as reported previously The ED10 and
doses of 1 or 2 Gy (but not 0.25 Gy) of 110-MeV protons de-
ED50, but not the ED90, values estimated for the fraction of animals
livered at a high dose rate of 0.5 Gy/minute or 0.25 and 1 Gy
that retched (or vomited) post-irradiation with protons at the HDR
(but not 2 Gy) of 110-MeV protons delivered at a low dose rate
were lower than the lower limits of the respective 95% confidence
of 0.5 Gy/hour Human PT values are com-
intervals previously reported for γ -rays suggesting
monly reported as an INR which is defined as the patient's ‘test'
that HDR proton irradiation was more effective than HDR γ -ray
PT value divided by the laboratory ‘normal' PT value, raised to
irradiation in inducing retching and vomiting.
the power of the International Sensitivity Index. INR values werecalculated for animals exposed to 2 Gy of 110-MeV protons at
2.4. Effects of radiation on blood coagulation and the development of
both the high and low dose rates and 1 Gy of 110-MeV protons
disseminated intravascular coagulation
at the low dose rate, which resulted in the greatest PT responseto proton radiation in the study. Three out of 10 animals ex-
Relatively little information exists in the literature on the ef-
posed to 1 Gy at the low dose rate had an INR value of � 2.0
fects of radiation on blood coagulation. Blood coagulation involves
and an additional 3/10 of the ferrets had borderline INR values
multiple components, which generate a fibrin-rich blood clot to
(> 1.75) approaching 2.0 which is con-
stop bleeding in a process known as hemostasis. Primary hemosta-
sidered to be of clinical concern for humans
sis starts with the activation of platelets at the wound site by
The INR values for the animals exposed to 2 Gy pro-
exposing collagen to blood, which allows von Willenbrand fac-
tons at the high dose rate were significantly higher than the pre-
tor (vWF) to bind to collagen and tethers platelets to the vas-
irradiation levels, although they were still below 2. In addition to
culature wall. The coagulation pathway, which is also referred
the increase in PT, aPPT also increased 3 and 48 hours after 0.25,
to as secondary hemostasis, occurs simultaneously on the nega-
1 or 2 Gy of 110-MeV proton irradiation at the low dose rate
tively charged surface of activated platelets to generate a fibrin-rich
For the ferrets irradiated with 110-MeV
thrombus The exposure of tissue
protons at the high dose rate, significant increases in aPPT was
factor (TF) at the injury site of blood vessels, and its subsequent
not observed in any of the radiation dose groups at 3 hours post-
binding with Factor VII (extrinsic pathway), initiates a thrombin
irradiation or in the 0.25 and 1 Gy dose groups at 48 hours post-
(Factor II) burst that leads to fibrin clot formation through the
irradiation. The increase in PT induced by the proton irradiation at
activation of a series of vitamin K-dependent serine proteases.
the high dose rate was due to Factor VII whereas Factors II, V, VII
It is the activation of coagulation factors, such as Factor V and
and IX contributed to the increases in PT induced by proton irra-
VIII, by thrombin that drives the development of a stable fibrin
diation at the low dose rate These results
clot as a part of the intrinsic pathway
demonstrated that proton irradiation significantly increased blood
clotting times due to different coagulation factors, indicating po-
Previous studies have demonstrated that radiation can induce
tential radiation-induced coagulopathy. The finding that the effects
vWF secretion from human umbilical vein endothelial cells irra-
of the proton radiation at the low dose-rate are more severe than
diated in tissue culture and the vWF mRNA
those at the high dose-rate on an endpoint is an unexpected find-
levels were increased when either human or bovine endothe-
ing in radiobiology, as the expectation is that reducing the dose
A.R. Kennedy / Life Sciences in Space Research 1 (2014) 10–43
rate will have a sparing effect, thereby reducing the severity of ef-
exposure to proton or γ -ray radiation produced step-wise changes
fect on the biological endpoint being evaluated. Thus, the increased
in hemostasis that begin with the radiation activated clotting cas-
effect of low dose-rate irradiation on ferret blood clotting times is
cade, which results in the cleavage of fibrinogen and the formation
of fibrin clots. Activation of the coagulation cascade also leads to
The blood clotting abnormalities in the ferrets are thought to
increased PT and aPTT values due to the consumption of associated
lead to a condition known as disseminated intravascular coagula-
factors These changes occurred as early as
tion (DIC), which is believed to result in 100% mortality in ferrets
3 hours post-irradiation, along with the detectable soluble fibrin in
irradiated with a 2 Gy dose of either γ -ray or SPE proton radiation
the blood as well as detectable fibrin clots in the blood vessels of
The increases in the blood clotting times
irradiated tissues In animals destined to die
became more evident in ferrets destined to die from exposure to
after irradiation, these abnormal hemostasis parameters became
radiation. The LD50 dose for the development of DIC in ferrets is
progressively worse. In ferrets exposed to a lower (sub-lethal) radi-
ation dose (e.g., 1 Gy), the abnormal hemostasis characteristics re-
While irradiation with SPE-like protons was shown to increase
covered steadily and by day-30 post-irradiation, the clot formation,
prothrombin time and partial thromboplastin time
clot size, and platelet clumping values returned to baseline. These
the mechanism for the proton radiation induced hypoco-
results are particularly important, since the recognition of radiation
agulation remains to be elucidated. We have hypothesized that
induced DIC as a cause of death could change the course of actions
the SPE-like proton irradiation activates the coagulation cascade,
when the acute radiation syndrome is diagnosed and treated in
which would put irradiated subjects in a hypocoagulable state. To
people exposed to radiation through occupational accidents, radia-
test this hypothesis, a separate experiment was performed with 12
tion terrorism or other catastrophic events
to 15 week old descented female ferrets irradiated with 1 Gy of
It is noteworthy that experiments performed as part of this
110-MeV protons at a dose rate of 0.5 Gy/hour, and the results in-
project also documented the development of DIC in Yucatan
dicate that the radiation exposure resulted in coagulation cascade
minipigs (Krigsfeld, G.S., Shah, J.B., Sanzari, J.K., Lin, L. and Kennedy,
activation, which was indicated by increases in soluble fibrin con-
A.R., Unpublished data). Three pigs were irradiated with a 2.5 Gy
centration in the blood and fibrin clots in blood vessels of livers,
homogeneous dose of total body x-irradiation. One of these pigs
lungs and kidneys from irradiated ferrets
died, and another was euthanized; both of these pigs were diag-
The soluble fibrin concentration was determined using a rapid sol-
nosed with DIC. The third pig did not die, but exhibited severe
uble fibrin assay that was previously developed and implemented
blood clotting abnormalities (greatly increased bleeding times,
at the Loma Linda University Medical Center to aid in early detec-
etc.), as did the two pigs that did not survive. In addition, three
tion of DIC in emergency room, operating room, or transplant pa-
pigs were irradiated with a 2 Gy total body dose of SPE-like proton
tients In addition to the activated coagulation
radiation. These pigs exhibited severe blood clotting abnormalities
cascade, PT and aPTT were also increased after irradiation, which is
(increased blood clotting times), but they survived the radiation
indicative of the involvement of the extrinsic/intrinsic coagulation
pathways. The platelet counts in the irradiated ferrets remained
2.5. Effects of radiation on fatigue
at approximately pre-irradiation values for up to 7 days post-irradiation, indicating that the observed effects on blood clotting
Several experiments have been performed to evaluate the abil-
times were not platelet-related. The activation of the coagulation
ity of SPE proton and γ -ray radiation to induce fatigue in mice,
cascade is expected to consume clotting factors, which, in turn,
as reflected in changes in social exploration, submaximal exer-
leaves the animal deficient in clotting factors. Thus, the increased
cise treadmill and locomotor activity
PT and aPTT values in the irradiated animals might have been due
The results of studies on social exploration indicated
to radiation-induced effects on secondary hemostasis. WBC counts
that low, but not high, dose-rate γ -ray and proton radiation expo-
were reduced significantly within 24 hours post-irradiation and
sures led to comparable transient increases in social withdrawal,
they remained reduced up to 7 days post-irradiation with a dose
and these effects are thought to be due to a combination of re-
of 1 Gy of SPE-like proton radiation.
straint stress and radiation. The results for studies on submaximal
DIC is a serious, life-threatening condition in which clotting
exercise treadmill indicated that neither γ -rays nor protons im-
and bleeding are occurring at the same time, and it is often fa-
paired performance on this test. In a study performed with 7 to
tal due to multiple organ failure. Mechanistically, activation of the
8-week old male CD-1 mice irradiated with 0.5 or 2 Gy 137Cs
clotting cascade is expected to decrease the bioavailability of the
γ -rays at a dose rate of 44.5 cGy/minute (high dose rate), 60Co
factors in the blood, thereby increasing the PT/aPTT values, as was
γ -rays at a dose rate of 0.5 cGy/minute (low dose rate) or protons
observed in the irradiated ferrets Ra-
at dose rates of 0.5 Gy/minute (high dose rate) or 0.5 cGy/minute
diation exposure may significantly decrease leukocyte counts, and
(low dose rate), locomotor activity was reduced in mice irradiated
the prolonged low WBC, neutrophil, and lymphocyte counts can
with γ -rays at the high dose rate but not in the mice irradiated
leave the irradiated subjects at risk for infection, thereby further
with γ -rays at the low dose rate or with protons, which had a
overwhelming hemostasis and potentially leading to DIC, as has
similar macroscopic dose distribution as that from 60Co γ -ray ex-
been observed in patients with sepsis. Currently, the mechanism
posure, when delivered at either the high or the low dose rate
of radiation-induced death at the dose expected to kill 50% of the
The γ -ray irradiation also increased hippocam-
irradiated subjects (LD50) is thought to be due to bone marrow cy-
pal TNF-α expression, which occurred as early as 4 hours post-
totoxicity (known as the hematopoietic syndrome), which results
irradiation and was followed by subsequent increases in IL-1RA in
in a dramatic reduction in the number of circulating hematopoietic
the cortex and hippocampus and reductions in activity-regulated
cells and the resultant symptoms of infection (from white blood
cytoskeleton-associated protein (Arc) in the cortex. These observa-
cell loss) and bleeding (presumably from platelet loss)
tions indicate that low-dose ionizing radiation rapidly activates the
However, studies performed with ferrets have sug-
neuroimmune system, potentially causing early onset fatigue-like
gested that the death of the animals irradiated at the LD50 dose is
symptoms in the irradiated animals.
due to a consumptive coagulopathy, which is followed by the on-
In these studies related to proton or photon induced fatigue,
set of DIC, since hypocoagulation occurred during early time points
the results were consistent with a threshold effect, i.e., once a
post-irradiation when the platelet counts were at normal levels
dose sufficient to produce a response is given, additional dose does
The ferret study results have shown that
not increase the magnitude of the response. The magnitude of the
A.R. Kennedy / Life Sciences in Space Research 1 (2014) 10–43
Fig. 7. Loss of blood vessels in the pig dermis after a high skin dose exposure to SPE-like radiation. Pig skin tissue samples were taken pre-irradiation (Panel A) and at 30
days after the pig was exposed to a 10 Gy skin dose of 6 + 12 MeV electron radiation (Panel B). For the skin tissue taken before irradiation, vascular beds are denoted by
squares (Panel A). A consequence of exposure to the high skin dose from SPE radiation is that blood vessels disappear in the areas beneath the epidermis (Panel B), such
that the blood flow to the cells in this area is reduced. Such areas of reduced blood supply beneath the epidermis occur in interventional radiology patients, and are known
to be life-threatening Thus, the areas with reduced blood supply must be removed surgically.
response with γ -rays is small, and additional stressors, such as
Denuded vasculature, isolated from untreated and irradiated ani-
restraint or other manual manipulations, are sufficient to obscure
mals, was unresponsive to BK treatment, confirming that the BK
any acute behavioral changes. For those tests in which altered
response is mediated via receptors present on the surface of en-
behavior was noted for γ -rays, protons showed a similar trend,
dothelial cells The fact that the relaxation
which did not reach statistical significance. Therefore, the RBE for
response was lower in irradiated tissue suggests that radiation ex-
proton radiation on this endpoint is assumed to be less than 1, al-
posure damages the vascular endothelium. The results of this study
though it could not be defined mathematically due to the lack of a
demonstrate that radiation has a direct effect on the cardiac vas-
statistically significant trend in the proton irradiated animals.
culature. Intact arteries from hearts taken from irradiated pigs ex-
The data gathered on the ability of SPE radiation to induce
hibited a reduced contractile force following exposure to BK. BK is
fatigue in mice suggest that exposure to low-dose rate ionizing
an endothelium-dependent dilator that acts directly on endothe-
radiation leads to a minimal increase in fatigue, in the form of
lial cells, causing them to release nitric oxide, PG12 and perhaps
depressive/anxious behaviors, and that these effects are transient
endothelium derived dilating factor(s), which signal relaxation of
with full recovery within the 24 hour period
vascular smooth muscle Our results suggest
These behaviors are likely of equal or lesser magni-
that alterations in vascular function are primarily a consequence
tude than the depressive/anxious behaviors that are stimulated by
of radiation-induced damage to endothelial cells, and that astro-
the restraint stress necessary to perform the experiments and that
nauts exposed to a high dose of SPE radiation could be at risk for
is likely to be experienced by astronauts in typical space explo-
vascular damage in the heart. While the skin dose was high in this
ration vehicles. Thus, acutely, SPE radiation up to a 2 Gy total body
study, the heart dose was only 0.35 Gy. This heart dose is consider-
dose is highly unlikely to increase fatigue or other adverse behav-
ably lower than the doses of radiation (0.5–5 G) previously shown
iors over and above baseline levels for astronauts and is therefore
to affect endothelial dependent relaxation in rat aorta
highly unlikely to lead to mission critical fatigue.
2.6. Heart functional changes
2.7. Skin effects
As part of the experiments designed to determine whether a
The skin effects in pigs have been described previously
high skin dose from SPE radiation has adverse effects on the in-
and the immunological changes in pig skin accom-
ternal organs of pigs, we evaluated changes in the heart brought
panying the skin changes are described above in the section on
about by a high skin dose of electron radiation planned to simulate
immunology effects. It has been observed that pigmentation in-
a dose distribution pattern like that expected from SPE radiation.
creases with increasing exposure to SPE radiation
As changes in the left anterior descending (LAD, interventricular)
and a method of quantitation for the melanin changes in
artery can have major effects on heart function, our studies in
irradiated pig skin has been developed (Billings, P.C., Sanzari, J.K.,
this area of research focused on LAD function in the pig hearts.
Kennedy, A.R., Cengel, A.K. and Seykora, J.T., Unpublished data). With
The LAD artery provides the blood supply to the mid-region of
very high skin SPE radiation doses, there is evidence of blood ves-
the heart and is a major site of vessel stenosis. Vessels from con-
sel loss in the pig dermis, as illustrated in A consequence
trol and pigs exposed to electron radiation exhibited a similar
of this radiation effect is reduced blood flow to these areas of the
relaxation response following treatment with adenosine diphos-
dermis. It is known that areas of the dermis receiving a reduced
phate and sodium nitroprusside. There was a reduced relaxation
blood flow can develop into a "diabetic foot" like state (which
response to bradykinin (BK) treatment in the arteries from hearts
needs surgical removal) in patients undergoing interventional radi-
exposed to SPE-like electron radiation. In contrast, vessels obtained
ology procedures In humans, the
from control animals exhibited a 20% higher relaxation response
development of such lesions requiring surgical removal takes ap-
with BK, compared with arteries obtained from the irradiated pigs.
proximately one year or more and the skin doses associated with
A.R. Kennedy / Life Sciences in Space Research 1 (2014) 10–43
such lesions are high (�10 Gy) Such lesions could
Upon evaluation of the optic nerve area, the meningeal sheath
conceivably be a problem during the multi-year exploration class
area was dilated in 31% of the irradiated animals, and the dif-
missions planned for the future.
ferences between the measured area of dilation in the irradiatedpigs compared to the non-irradiated pigs were statistically sig-
2.8. Increased intracranial pressure and effects potentially related to
nificant. Further, an accumulation of lymphocytes, plasma cells,
vision abnormalities evaluated in Yucatan mini-pigs
and macrophages around the vessels near the optic nerve wasobserved, indicating an inflammatory response to radiation expo-
Vision changes, characterized as a degradation in distant and
sure; however, the presence of inflammatory infiltrates was not
near visual acuity, have been documented in numerous astronauts
consistent in the irradiated animals (inflammatory infiltrates were
who have been involved in long-duration (of six months or longer)
present in 10% of the irradiated pigs).
space flight Although the etiology of the vi-
The results described above suggest that SPE radiation may re-
sion alterations is unknown, Mader et al. hypothesized that the
sult in radiation-induced retinal atrophy or degeneration; however,
optic nerve and ocular changes observed in astronauts could have
long term studies will be necessary to determine whether the loss
been caused by prolonged microgravity exposure
of photoreceptor cells and changes in retinal width or optic nerve
Since radiation exposure in clinical radiotherapy patients
area persist at later time periods.
is known to be associated with increased intracranial pressure,increased radiation exposures during space travel could also con-
2.8.2. Ocular ultrasound results of the eyes of the pigs exposed to SPE
tribute to the alterations thought to be involved in producing
the vision abnormalities. We have evaluated the ability of SPE-
Noninvasive measurements such as those obtained from bed-
like radiation in pigs to produce changes like those associated
side ocular ultrasound examinations have been advocated and uti-
with the astronaut vision alterations, by performing ocular ultra-
lized in the clinical setting and during spaceflight to give informa-
sound examinations, lumbar puncture opening pressure studies
tion about changes potentially related to vision abnormalities. Live
and histopathologic examinations of eye tissue taken from pigs ir-
ocular ultrasound imaging was performed in the pSPE animals ap-
radiated with SPE-like radiation (in the forms of simulated electron
proximately 2 months post-irradiation as part of a collaborative
(6 + 12 MeV) SPE (eSPE) or simulated proton SPE (pSPE) exposures
study between the investigators at the University of Pennsylvania
The dosimetry studies have indicated that
(Penn) and Wyle/NASA investigators skilled in ocular ultrasound
the eyes and lenses receive substantial doses of radiation during
examination techniques. The sonographer at Penn was remotely-
an SPE; the dose to the lens is roughly comparable to the dose
guided by an expert in eye/optic nerve imaging, and the remotely
received by the skin of pigs (e.g., see
guided sessions were conducted in a way that was similar to the"telemedicine" and "telescience" arrangement routinely used dur-
2.8.1. Histopathology changes in the eyes of pigs exposed to simulated
ing ultrasound imaging sessions on the International Space Station
proton and electron SPE radiation which could be related to vision
(ISS). The optic nerve sheath diameter was measured using elec-
tronic calipurs at three distances from the vitreoretinal interface.
The retina is a layered structure of neurons interconnected by
Animals exposed to 5–10 Gy pSPE radiation exhibited a dose-
synapses sending information to the brain via the optic nerve. The
dependent trend in increased diameter measurements compared
retina includes millions of photoreceptor cells, known as rods or
to the non-irradiated animals. The measurements recorded for the
cones, which are sensitive to light. Change or damage to the retina
animals exposed to 5 Gy pSPE radiation resulted in a significant
can cause loss of vision. Signs of damage to the retina are sudden
increase (p = 0.02 for right eye; p = 0.05 for left eye), compared
flashes of light, floating spots, decreased vision, or distorted vision.
to the non-irradiated group of animals. The 10 Gy dose pSPE ra-
The ocular histopathology of pigs exposed to pSPE (5–10 Gy) or
diation resulted in significantly increased sheath diameters as well
eSPE (5–20 Gy) radiation was examined. In this study, (Sanzari, J.K.,
(p = 0.002 for right eye; p = 0.03 for left eye) (Sanzari, J.K., Sar-
Zeiss C.J., and Kennedy, A.R., Unpublished data) sagittal sections of
gysan, A.E., Ebert, D., Garcia, K.M., Shultz, S.M., Seghal, C.M., and
the entire eye and cross sections of the optic nerve were prepared
Kennedy, A.R., Unpublished data). Ocular ultrasound examinations
and qualitative differences (as well as some quantitative measure-
were not performed on the corresponding animals exposed to eSPE
ments) between the eyes of the irradiated and non-irradiated ani-
radiation (for which histological changes are reported above).
mals were investigated.
When considered together, the histopathology and the ocular
It was determined that eSPE radiation exposure resulted in a
ultrasound exam results indicate that SPE radiation exposure di-
decreased total retinal width in 20% of the irradiated animals com-
rectly affects the eye structure of animals exposed to 5–20 Gy
pared to the non-irradiated control animals; the differences ob-
electron simulated SPE radiation or 5–10 Gy proton simulated SPE
served in these studies between the irradiated and control pigs
radiation. The observed changes in the optic nerve area may be
were statistically significant. Changes in retinal width can cause
associated with the optic disc edema observed during/post space-
turbulence in blood flow and may indicate atrophy affecting the
flight reported by
nerve fiber layer, ganglion cell layer, outer plexiform layer, and in-ner nuclear layer of the retina. It was confirmed that the width of
2.8.3. Opening pressure in pigs exposed to proton and electron SPE
the outer plexiform layer was also reduced in some of the animals
exposed to eSPE radiation. A reduction in the width of the outer
The opening pressure of pigs exposed to either pSPE or eSPE
nuclear layer is an indicator of photoreceptor cell loss, which was
was measured by lumbar puncture procedures and was found to
confirmed by the extrusion of photoreceptor nuclei in the retina.
be increased in some of the irradiated animals, with relatively
The extrusion of photoreceptor nuclei (an indication of active cell
larger increases in animals exposed to larger electron or proton SPE
death) into the inner segments was observed in both the eSPE and
radiation doses Pigs exposed to lower skin
pSPE animals. The dose–response relationship was evaluated for
doses of 2.5 to 7.5 Gy eSPE radiation (like the SPE radiation occur-
the extrusion of photoreceptor nuclei induced by pSPE radiation
ring in October 1972) exhibited increased opening pressure values,
exposure and results indicated a statistically significant slope, es-
which lasted up to 90 days post-irradiation (i.e., at the time when
tablishing that the loss of photoreceptor nuclei in pSPE irradiated
the experiment was terminated), suggesting that SPE-like electron
pigs was dose-dependent.
radiation resulted in increased intracranial pressure after a radia-
A.R. Kennedy / Life Sciences in Space Research 1 (2014) 10–43
tion exposure with a skin dose as low as 2.5 Gy
to 3 Gy protons (p < 0.001) or 0.5 Gy 56Fe ions (p < 0.05) than
in mice receiving only the sham radiation
The results in this area of research indicate that exposure to
The decreased survival in the irradiated mice was accompanied
even relatively low skin doses of SPE radiation can result in some
by a significant increase in the rate of development of malignant
of the alterations thought to play a role in astronaut vision alter-
lymphoma and Harderian gland tumors as well as the fractions of
ations (e.g., increased intracranial pressure and increases in nerve
animals with malignant lymphoma or rare tumors
sheath diameter). Thus far, astronauts have not been exposed to
significant doses of SPE radiation during spaceflight, but it is ex-pected that there will be an increased risk of astronaut exposure to
3.2. Cataract development
higher doses of SPE radiation in the exploration class missions ofthe future. It is hypothesized that exposure to SPE radiation along
In the long-term experiment with CBA/JCR HSD mice exposed
with extended space travel could exacerbate the development of
to 1-GeV protons at a dose of 3 Gy or 1-GeV/n 56Fe ion radiation
visual changes in astronauts. However, larger numbers of exposed
at a dose of 0.5 Gy, the mice were observed daily over approxi-
pigs than those used in the studies described above will be neces-
mately two years after the radiation exposure. The animals were
sary to confirm and verify this hypothesis.
then euthanized and lenses were harvested and characterized us-ing an established classification system that assigns discrete scores
2.9. Short-term survival in irradiated animals
based on the severity of the lens opacifications. The results showedthat exposure to 1-GeV/n proton (3 Gy) or 56Fe ion (0.5 Gy) radi-
In experiments performed with male ICR mice aged 4 to 5
ation significantly increased the cataract prevalence and severity
weeks exposed to 5.9, 6.8 or 7.2 Gy of total body irradiation with
in CBA/JCR HSD mice to levels above the baseline levels of age-
1-GeV protons at dose rates ranging from 0.2 to 0.7 Gy/minute,
induced cataract formation in this mouse strain
the 30-day survival was 60%, 13.3% and 0%, respectively, and thecalculated lethal dose to kill 50% of the irradiated animals was
3.3. Cancer development
6.23 Gy In a separate experiment performedwith male ICR mice aged 4 to 5 weeks irradiated with 6 or 8 Gy
Malignancy is considered to be a particular risk associated with
of 225 kVp X-rays, the 30-day survival was 100% and 6.7%, respec-
exposure to the types of ionizing radiation encountered during
tively these survival levels are higher than
space flight. In these studies, the ability of protons and highly
those observed in the 30-day survival studies in the animals ex-
energetic, heavy particles (HZE particles) to induce carcinogene-
posed to 5.9–7.2 Gy of 1-GeV/n protons. These results indicate that
sis was determined in CBA mice. The major finding of the studies
1-GeV proton radiation is more lethal to mice than X-rays. Similar
was that there was an increased risk of developing malignant lym-
studies were performed by other investigators in C57BL/6J mice to
phoma and rare tumor types, including Harderian gland tumors, in
determine the relative toxicity of HZE radiation (1-GeV/n 56Fe ions)
animals exposed to space radiations
compared to γ -rays or 1-GeV protons In these
A significant increase in pre-malignant and malignant lesions of
studies, the LD50/30 values for 56Fe ions, protons and γ -rays were
myeloid origin was also observed in mice exposed to 3 Gy pro-
reported to be 5.8, 6.8 and 7.25, respectively; the RBE value for the
ton radiation and 0.5 Gy 56Fe ion radiation. These results indicate
56Fe ions was 1.25 and for the protons, the RBE was 1.06. It was
that exposure to space radiations can increase the incidence of
concluded from these studies that 56Fe ions caused accelerated and
malignant tumors as well as pre-malignant lesions in mice. These
more severe hematopoietic toxicity. Of interest in this study was
studies have been reviewed recently
the finding that intestinal crypt cells did not show increased HZEtoxicity. In another reported study performed in C57BL/6J mice, it
4. Radiation induced changes in gene expression
was observed that the LD50/30 values for 28Si and 12C ions were5.17 and 7.34 Gy, respectively In these stud-
In a study performed with 5–6 weeks old ICR mice, irradia-
ies, the RBE values for 28Si and 12C ions (compared to the γ -ray
tion with 1-GeV protons was shown to increase the mRNA levels
data in which the LD50/30 was 7.25) were 1.4 and 0.99, respec-
for Bax, caspase-9, caspase-8, NFκ B1 and TGFβ1 and protein levels
for Bcl2 and Bcl-xL The proton irradiation
In studies performed with ferrets, the observed 30-day survival
was also shown to induce cleavage of pro-apoptosis proteins, such
was 100% for ferrets irradiated with up to a 1 Gy dose of SPE-like
as caspase-3 and PARP-1, in bone marrow lysates of the irradiated
protons and zero for ferrets exposed to 2 Gy of SPE-like protons
animals. These results confirm the findings that the high-energy
The LD50/30 for SPE-like proton radiation
proton radiation can induce the gene expression of classical mark-
in ferrets was estimated to be approximately 1.5 Gy
ers of apoptosis, as well as the downstream effectors, caspase-3
The survival curves for γ -ray irradiated ferrets were
and PARP-1. In a separate study performed with ICR mice irradi-
comparable to those for the proton-irradiated animals, and both
ated with 1.0 and 6.4 Gy of 1-GeV protons or 1.1 and 7.0 Gy of
the proton and γ -ray irradiated ferrets displayed signs of distress
γ -rays to compare the acute effects of radiation on gene expres-
including ecchymosis, petechiae, and hemorrhaging. It was hypoth-
sion in radiation-sensitive tissues (e.g., spleen, thymus, bone mar-
esized that the ferrets were dying of DIC in these studies
row, testis and the GI tract), the apoptotic responses were found
which was confirmed by additional evidence
to vary greatly between γ -ray and proton irradiated animals in a
tissue- and dose-dependent manner and cell death in the splenicwhite pulp was consistently lower in the proton-irradiated animals
3. Long-term effects
compared to the γ -ray irradiated animals Both proton and γ -ray irradiation triggered nuclear accumulation
3.1. Long-term survival in irradiated animals
of p53, with no significant differences in the majority of the knownpro-apoptotic p53-target genes in the spleens of irradiated mice.
In a long-term study in which male CBA/JCR HSD mice aged 8
However, γ -ray irradiation uniquely triggered a pro-apoptotic ex-
to 9 weeks were irradiated with 3 Gy of 1-GeV protons or 0.5 Gy
pression profile in the spleen and Peyer's patches, which exhibited
of 1-GeV/n 56Fe ions and monitored for 2 years after irradiation,
a higher level of apoptosis after γ -ray irradiation than after the
the survival of the mice was significantly lower in mice exposed
proton irradiation despite the increased presence of DNA strand
A.R. Kennedy / Life Sciences in Space Research 1 (2014) 10–43
breaks and phosphorylated-ATM in the spleens of proton irradi-
cell membrane components, antioxidants could be effective coun-
ated animals. Differences in the acute pro-apoptotic response to
termeasures against radiation induced oxidative stress and other
proton and γ -ray irradiation correlated with increased expression
adverse biological effects occurring downstream to the free radical
of the p53-dependent pro-apoptotic gene, Bcl-G, and granzyme B,
suggesting that the fate of the cells after proton and γ -ray irra-
The agents evaluated in our studies as potential countermea-
diation may be context-dependent and the triggering of apoptosis
sures for radiation induced oxidative stress included N-acetyl cys-
in lymphoid cells after irradiation may not be dependent solely on
teine (NAC), ascorbic acid (or vitamin C), coenzyme Q10, folic acid,
the extent of DNA damage brought about by the radiation expo-
glutathione, α-lipoic acid, niacin, L-selenomethionine (SeM), thi-
amin and vitamin E succinate. NAC is a small molecular weight
In female ICR mice irradiated with 60Co γ -rays at low
thiol and a precursor to intracellular cysteine and glutathione
(0.5 Gy/hour) or high (0.5 Gy/minute) dose rates, changes in the
which is a tripeptide small molecular weight thiol
expression of genes implicated in oxidative stress, extracellular
shown to be a versatile protector against radiation induced ox-
matrix (ECM) remodeling and selected protein expression profiles
idative damage NAC is also effective
in mouse skin were examined using skin tissues harvested at 4
in activating NF-κB and manganese superoxide dismutase (Mn-
hours post-irradiation After irradiation at doses
SOD) gene expression
as low as 0.25 Gy, the expression of many genes responsible for
MnSOD is a main mitochondrial antioxidant enzyme with ra-
regulating the production of reactive oxygen species were signif-
dioprotective properties
icantly altered by more than 2-fold as compared to unirradiated
Vitamin C is a water-soluble antioxidant that
controls. The expression profiles of 18 to 20 of the 84 ECM genes
reacts with highly damaging hydroxyl radicals to form less toxic
were also significantly altered after irradiation at the low dose
ascorbate free radicals, which can be detoxified by enzymes that
rate. As compared to the low dose rate irradiation, the high dose
reduce ascorbate free radicals back to ascorbic acid
rate irradiation resulted in different ECM gene expression profiles,
Vitamin E is a lipophilic agent that protects cell membranes from
with the most striking differences observed for genes encoding
oxidative damage by radiation or other physical or chemical agents
matrix metalloproteinases. These results indicate that the expres-
Dietary supplementation of vitamins C and E
sion of many genes involved in oxidative stress responses and ECM
is thought to be important for protection against human diseases
remodeling may be differentially regulated by high and low dose
associated with free radical damage to cellular DNA, lipids and pro-
rate irradiation.
teins Lipoic acid is a B vitamin that is bothlipid and water-soluble and is considered to be a "universal antiox-
5. Countermeasures and mitigation of space radiation damage
idant" because it can react with hydroxy radicals, singlet oxygen,and peroxyl and hypochlorous radicals It is anexcellent radical scavenger both in the oxidized and reduced form
It has been well established that the biological effectiveness of
and is known to regenerate other antioxidants from their inactive
ionizing radiation depends on the LET, which describes the rate of
forms. As examples, lipoic acid plays an essential role in mito-
energy loss along the trajectory of ionizing particles
chondrial dehydrogenase reactions It also protects
and the ion species
cell membranes by reacting with and regenerating vitamin C and
Ionizing radiation damages cells through a combination of direct
glutathione, which in turn recycle vitamin E
action, which refers to the direct hit of biologically important
Treatment with lipoic acid has been shown to reduce radiation-
targets by the particle radiations, and indirect actions via water-
induced oxidative stress
derived free radicals produced by the radiation
and hematopoietic tissue damage in irradi-
While the indirect action plays an important role in the bi-
ated mice Treatment with lipoic acid
ological effects of low-LET radiations, such as X-rays and γ -rays,
in combination with vitamins C and E has been shown to pro-
its contribution diminishes with increase in LET and the direct ac-
tect against lens damage caused by low dose irradiation
tion contributes more to the biological effects of high-LET radiation
Selenium is an essential trace element for main-
than the indirect action
taining activities of the important antioxidant enzymes, thiore-
From our studies, there have been many publications related
doxin reductase and glutathione peroxidase
to countermeasures for acute radiation effects
Together with vitamin E, it pro-
tects cell and organelle membranes from oxidative damage, facil-
itates the union between oxygen and hydrogen at the end of the
metabolic chain and the transfer of ions across cell membranes
These publications are
Another agent evaluated as a potential
briefly described below.
countermeasure for radiation induced oxidative stress is a soybean-derived protease inhibitor known as the Bowman–Birk inhibitor
5.1. Radiation induced oxidative stress and antioxidants as
(BBI), which has been developed in the form of BBI Concentrate
(BBIC) for cancer prevention and human trials, as reviewed pre-viously Both BBI and BBIC
Using a dichlorofluorescein (DCF) fluorometric assay that we
have been shown to have antioxidant properties
had previously adapted and standardized to measure radiation in-
and BBI has been utilized as a radioprotective agent
duced oxidation in live cells
we have demonstrated that low LET photon radiation, such as X-
rays and γ -rays, and high LET radiation, such as 0.6-GeV/n silicon
Using the DCF fluorometric assay method previously adapted
ions and 1- or 5-GeV/n 56Fe ions, as well as 250 MeV protons, are
for measurement of radiation induced oxidative stress in cultured
all capable of inducing oxidative stress sug-
cells we performed experiments with X-
gesting that the indirect actions via free radical generation may
rays, γ -rays, protons and HZE particles to evaluate the protective
contribute substantially to the biological effects of both low and
effects of antioxidants against radiation induced oxidative stress
high LET radiation. Since the removal of radiation generated free
and found NAC, ascorbic acid, α-lipoic acid and SeM to be highly
radicals will make them unavailable to damage DNA, proteins and
effective in preventing radiation induced oxidative stress, whereas
A.R. Kennedy / Life Sciences in Space Research 1 (2014) 10–43
coenzyme Q10 and vitamin E succinate were only weakly effective
determined in HTori-3 cells by the soft agar colony formation as-
in preventing radiation induced oxidative stress in cultured cells
say, which measures the capability of HTori-3 cells to grow in
Based on the results of
an anchorage-independent manner. The results indicate that treat-
the DCF fluorometric assay experiments, these antioxidants were
ment of the cells with BBI, BBIC, SeM alone or in combination with
selected as a combination for further studies in vitro and in vivo.
ascorbic acid, coenzyme Q10 and vitamin E succinate prevented
The ability of the antioxidant combination and BBIC to pre-
proton and HZE particle radiation induced HTori-3 cell transforma-
vent radiation induced oxidative stress in vivo was evaluated in
tion in vitro In experiments performed
Sprague-Dawley rats and CBA mice irradiated with γ -ray, proton or
with γ -ray radiation, treatment with 5 μM SeM before, during
56Fe ion radiation using the total antioxidant status (TAS) in serum
and/or as late as 7 days after the radiation exposure brought the
or plasma as the biological endpoint
anchorage-independent colony formation efficiency down to levels
In the rat studies, the serum or plasma
that were not significantly different from the sham radiation con-
level of total antioxidants was found to be decreased after expo-
sure to γ -ray or 1-GeV/n 56Fe ion radiation, and the decrease wasalleviated or completely prevented in the animals fed with a diet
5.3. Antioxidant protection against space radiation induced mortality
supplemented with SeM (12 μg/g diet) alone or in combinationwith sodium ascorbate (19 μg/g diet), NAC (51 μg/g diet), α-lipoic
It is well known that the hematopoietic system is highly sen-
acid, reduced form (100 μg/g diet), vitamin E succinate (8.6 μg/g
sitive to total body irradiation (TBI) and the fate of hematopoietic
diet) and coenzyme Q10 (51 μg/g diet)
cells after TBI may determine the survival or death of irradiated
In the mouse studies, the plasma TAS also
subjects Thus, we
decreased significantly after exposure to 0.5 Gy of 1-GeV/n 56Fe
have evaluated the effects of antioxidants in mice using the 30-day
ion radiation or 3 Gy of 1-GeV proton or γ -ray radiation, and
survival level and hematopoietic cell counts as the biological end-
the decrease in plasma TAS was alleviated or prevented com-
points. Dietary supplementation with an antioxidant combination
pletely by diet supplementation with BBIC (10 mg/g diet), SeM
consisting of SeM (0.06 μg/g diet), α-lipoic acid (85.7 μg/g diet),
(0.14 μg/g diet), or a combination of L-SeM (0.14 μg/g diet),
NAC (171.4 μg/g diet), sodium ascorbate (142.8 μg/g diet) and vita-
sodium ascorbate (17.14 μg/g diet), NAC (51.43 μg/g diet), α-lipoic
min E succinate (71.4 μg/g diet) significantly improved the 30-day
acid (102.86 μg/g diet), vitamin E succinate (8.57 μg/g diet) with
survival of the mice irradiated with 8 Gy of X-rays
or without coenzyme Q10 (51.43 μg/g diet)
or 5.9 Gy of protons However, no sig-
These results indicate that BBIC, SeM and
nificant improvement was observed for the mice irradiated with
the antioxidant combinations are potentially useful as countermea-
protons at higher radiation doses (6.8 or 7.2 Gy)
sures against space radiation-induced oxidative stress and subse-
It is expected that the higher proton radiation doses used
quent adverse biological effects, which could arise from the in-
in these studies might have caused damage that was beyond miti-
creased oxidative stress in irradiated subjects.
gation by the antioxidant treatment. In both the proton and X-rayradiation experiments, antioxidants were more protective when
5.2. Antioxidant protection against radiation induced cell death and
the antioxidant treatment was initiated 2 hours after radiation ex-
transformation in vitro
posure as compared to the antioxidant treatment initiated 7 daysprior to radiation exposure These re-
The protective effects of the antioxidants, BBI and BBIC against
sults were confirmed by other investigators, who also showed that
radiation induced cell death have been evaluated in vitro using
the antioxidant combination had a better protective effect on radi-
the clonogenic survival of cultured MCF10 cells or HTori-3 cells
ation induced lethality when started 7 days post-irradiation than
as the biological endpoints Irradi-
it did when it was started 2 hours post-irradiation
ation with 5-GeV/n 56Fe ions resulted in a dose-dependent de-
The findings that antioxidant treatment leads to better sur-
crease in the clonogenic survival of the MCF10 cells, which was
vival when applied post-irradiation might result from an adaptive
attenuated by treatment with SeM alone or in combination with
response to radiation exposure, which has been reviewed recently
ascorbic acid, coenzyme Q10 and vitamin E succinate with an es-
The ability of the antioxidants to
timated dose modifying factor (DMF) of 1.2, 1.4, 2.2 and 2.2, re-
improve radiation survival even when the antioxidant treatment
spectively, for BBI, BBIC, SeM alone or in combination with other
was initiated after the radiation exposure suggests that antiox-
antioxidants The identical DMF values for
idants can be a feasible countermeasure for radiation exposure
treatments with SeM alone or in combination with other antioxi-
associated with space travel, radiation accidents or terrorist attacks
dants indicate that these antioxidant combinations were not more
in which radiation exposure could occur without much advanced
effective than the SeM treatment alone under the experimental
conditions utilized. These results do not rule out the possibil-
The antioxidant treatment significantly attenuated the radia-
ity that the antioxidant combinations might have provided bet-
tion effects on peripheral hematopoietic cell counts in the mice
ter protection than SeM treatment alone under other experimen-
irradiated with 1 or 8 Gy of X-rays or 1
tal conditions or for other biological endpoints, however, since
or 7.2 Gy of 1-GeV protons The antioxi-
a combination of antioxidants with different biochemical proper-
dant treatment was also shown to improve the recovery of the
ties and action mechanisms is likely to provide better protection
bone marrow cell counts in mice irradiated with γ -rays
for different molecular targets during and after irradiation. For
or 1-GeV protons Thus, an-
examples, cationic thiols are much more effective than anionic
tioxidants appear to be effective for protection of hematopoietic
thiols in protecting DNA against radiation damage
cells against the adverse effects of either photon or proton radi-
and lipophilic antioxidants, such as vitamin
ation. The ability of antioxidants to prevent the radiation caused
E, are effective protectors of biomembranes
loss of circulating neutrophils, also called PMNs, are illustrated in
whereas hydrophilic antioxidants are more ef-
which shows that the mice maintained on an antioxidant
fective in protecting soluble proteins and enzymes in the aqueous
diet and exposed to an 8 Gy dose of radiation experienced a drop
environment of cells
in the levels of PMNs/neutrophils of a magnitude comparable to
The protective effects of the antioxidants, BBI and BBIC, against
the effects observed for mice exposed to a 1 Gy dose of radia-
proton and HZE particle radiation induced cell transformation were
tion and maintained on the normal diets. The mice maintained
A.R. Kennedy / Life Sciences in Space Research 1 (2014) 10–43
or 0.5 Gy of 1-GeV/n 56Fe ions, treatment with the antioxidantcombination or BBIC decreased the fractions of animals with ma-lignant lymphoma to levels that were not significantly differentfrom the baseline level The treatment withthe antioxidant combination or BBIC also prevented the increasein the fractions of mice with premalignant or malignant lesionsof myeloid origin after the proton irradiation and the incidencerate of rare tumors (which included Harderian gland tumors) af-ter the proton or 56Fe ion radiation exposures From these experiments, it was concluded that an-tioxidants have a major protective effect against space radiationinduced carcinogenesis in vivo Inthese studies, a major protective effect resulted from the abilityof the antioxidants to prevent the early stage neoplastic growthsfrom growing into fully developed, malignant tumors. Other stud-ies suggest that anticarcinogenic agents can be added at late timesfollowing carcinogen exposure, in both in vitro and in vivo sys-
Fig. 8. Effects of radiation on polymorphonuclear (PMN)/neutrophil cell counts in
tems, and still have a suppressive effect on the carcinogenic pro-
mice treated with antioxidants. In this experiment, animals were maintained on the
cess The results of the studies on radiation
control diet or antioxidant supplemented diet and irradiated with x-rays at doses of
induced carcinogenesis suggest that antioxidant and BBIC supple-
1 or 8 Gy, which significantly decreased the numbers of circulating PMN cells (neu-trophils) to a greater extent in mice maintained on the control diet than in mice
ments could be useful for the prevention of malignancies and other
maintained on the antioxidant supplemented diet. The magnitude of PMN cell loss
neoplastic lesions developing as a result of exposure to space radi-
in mice maintained on the antioxidant diets and exposed to 8 Gy of radiation was
ation. It has been concluded from these studies that antioxidants
comparable to that observed in the mice maintained on the control diet and ex-
have a major protective effect against radiation induced carcino-
posed to a 1 Gy dose of radiation. The decrease in the PMN cell counts in micemaintained on the antioxidant supplemented diet and exposed to a 1 Gy dose of
genesis, and are effective even when added at late times during
radiation was not significantly different from that in the control animals. These re-
the carcinogenic process.
sults indicate major beneficial effects of antioxidants on the survival of circulatingPMN cells/neutrophils following the radiation exposure. [These data are representa-
5.6. Mechanism(s) for antioxidants as radiation countermeasures
tive of those published previously
To study the mechanisms for the radioprotection by the an-
on the antioxidant diets and exposed to a 1 Gy dose of radia-
tioxidant treatment, we have examined the expression of the ATR
tion had PMN/neutrophil cell counts that were not significantly
gene, which is one of the central components of the DNA dam-
different from those in the control animals. Thus, it is concluded
age response pathway and the CHK2
that the antioxidant diet had highly significant beneficial effects on
gene, which is a cell cycle checkpoint regulator and putative tumor
PMN/neutrophil cell counts in irradiated animals.
suppressor in HTori-3 cells ir-radiated with 0.4 Gy of 5-GeV/n 56Fe ions with or without SeM
5.4. Antioxidant protection against space radiation induced cataracts
(5 μM) treatment initiated 24 hours prior to the radiation expo-sure. The results indicate that the ATR mRNA level was increased
An increased rate of cataract formation has been observed in
by 42% with the SeM treatment alone and increased by 94% with
astronauts which has
the SeM treatment and radiation exposure
been attributed to the increased exposure to cosmic radiation dur-
In the same study, SeM treatment alone did not significantly affect
ing space travel. To investigate the ability of BBIC and antioxidants
the CHK2 mRNA level, but the combined treatment with SeM and
to reduce the formation and severity of cataracts related to space
56Fe ion radiation increased the level of CHK2 mRNA by 99%. The
radiation exposure, mice were exposed to 1-GeV proton or 1-GeV/n
up-regulation of ATR and CHK2 gene expression observed in the
56Fe ion radiation and fed with a control diet or diets supple-
irradiated HTori-3 cells may prevent the cells from going through
mented with BBIC or the antioxidant combination containing SeM,
mitosis until the damage is repaired, thereby preventing the ra-
NAC, ascorbic acid, coenzyme Q10, α-lipoic acid and vitamin E suc-
diation damage from being fixed and leading to mutations and/or
cinate before and for 2 years after the radiation exposure
Lenses were harvested approximately 2 years after
In ICR mice irradiated with 6.4 Gy of 1-GeV protons or 7.0 Gy
the radiation exposure for evaluation of the lens opacifications. The
of γ -rays, 15 genes belonging to the class of "apoptosis regula-
results showed that treatment with BBIC or the antioxidant com-
tor activity" were differentially expressed in the spleen of mice fed
bination decreased the prevalence and severity of the lens opacifi-
an antioxidant supplemented diet as compared with mice fed with
cations in the mice irradiated with 3 Gy of 1-GeV proton or 0.5 Gy
the control diet, and the antioxidant treatment inhibited apoptosis
of 1-GeV/n 56Fe ion radiation, although statistical significance was
in the white pulp of the spleen following γ -ray irradiation, pos-
only achieved for the 56Fe ion irradiated mice, possibly due to the
sibly by altering IL-6 signaling and by blocking the expression of
higher proton radiation dose (3 Gy) that might have exceeded the
the prokineticin PROK2, the ligand to the G protein-coupled re-
protective capacity of the antioxidant combination or BBIC. These
ceptors PROKR1 and PROKR2 which are
results indicate that BBIC and the antioxidant combination could
involved in a number of pathophysiological processes. In ICR mice
be useful for protecting astronauts against space radiation-induced
irradiated with 1 or 8 Gy of X-rays, bcl-2 gene expression was
cataracts during or after long-term manned space missions.
found to be decreased, whereas bax, caspase 7, caspase 9 andTGF-β1 gene expression was increased at 4 or 24 hours after irra-
5.5. Antioxidant prevention of space radiation induced cancer
diation. Dietary supplementation with antioxidants attenuated theradiation effects on bax, caspase 7, caspase 9 and TGF-β1 gene
Radiation induced malignancy is a particularly important risk
expression, but increased bcl-2 gene expression by 10 fold at 24
associated with extended space travel. In a 2-year study performed
hours after irradiation The abrogated pro-
with CBA/JCR HSD mice irradiated with 3 Gy of 1-GeV protons
apoptosis (bax and caspase 9) gene expression and the increased
A.R. Kennedy / Life Sciences in Space Research 1 (2014) 10–43
anti-apoptosis (bcl-2) gene expression observed in the irradiated
experiment performed with ferrets (aged 12–15 weeks), that were
animals treated with antioxidants have implicated apoptosis as a
exposed to a 2 Gy total body dose of γ -ray radiation with sub-
key process modulated by antioxidants to attenuate the effects of
cutaneous injection of phosphate buffered saline (PBS) or Neulasta
radiation on the hematopoietic system and animal survival
(0.1 mg/kg) on days 1, 4, and 7 post-irradiation, peripheral blood
The anti-apoptotic effects of selenium have also been
was collected from each animal on day 0 (prior to radiation ex-
reported in normal human or mouse fibroblasts
posure), and on days 1, 4, 7, and 13 post-irradiation for analysis.
and primary human keratinocytes irradiated
The results demonstrate that the Neulasta treatment of irradiated
with ultraviolet irradiation. The anti-apoptotic effect of selenium
ferrets could lead to a significant increase in neutrophil counts
observed in normal animals or tissues after irradiation is in con-
(Krigsfeld, G.S., Sanzari, J.K. and Kennedy, A.R., Unpublished data). In a
trast to the pro-apoptotic effect of selenium observed in malignant
similar experiment performed with pigs (aged 12–15 weeks) that
cell lines or tissues
were exposed to a 2 Gy total body dose of SPE-like proton radi-
The differential effects of selenium on apop-
ation and subcutaneous injections of PBS or Neulasta (0.1 mg/kg)
tosis in normal and malignant cells/tissues suggest the possibility
on days 4, 7, and 10 post-irradiation, peripheral blood was col-
that selenium may protect normal tissues against radiation dam-
lected from each animal on day 0 (prior to radiation exposure),
age without the unintended consequence of suppressing radiation
and on days 1, 7, 10, 13, and 30 post-radiation for analysis. The
induced apoptosis in malignant cells or tissues. The different ef-
results indicated that the proton radiation exposure led to statisti-
fects of antioxidants in normal cells, as compared to their effects
cally significant decreases in neutrophil counts at days 7, 10 and
in malignant cells, or those in different stages of carcinogenesis,
13 post-irradiation in the irradiated animals injected with PBS;
have been discussed elsewhere
however, in irradiated pigs treated with Neulasta, the neutrophil
In a separate study performed with cultured HTori-3 cells ex-
counts were never decreased to below the baseline levels (Sanzari,
posed to low doses (0.1 and 0.2 Gy) of 1-GeV/n 56Fe ion radiation,
J.K., Krigsfeld, G.S., Shuman, A.L. and Kennedy, A.R., Unpublished data).
treatment with 5 μM SeM in the medium for 24 hours prior to ir-
The overall conclusion from the studies in mice, ferrets and pigs is
radiation profoundly affected the radiation induced alterations in
that Neulasta can increase the number of circulating neutrophils in
gene expression The exposure to 0.1 and 0.2
three different species of animals evaluated.
Gy of 56Fe ion radiation induced significant differential expressionof 196 and 610 genes, respectively, and the differential expression
5.8. SI–Wu–Tang or fructose as countermeasures to increase neutrophil
of 39% to 55% of these genes was abolished by the SeM treat-
counts in mice exposed to SPE or γ -ray radiation
ment Genes and functional pathways thatwere significantly up- or down-regulated by the 56Fe ion irradi-
A Chinese formula SI–Wu–Tang (SWT,
ation in the absence, but not in presence, of SeM treatment have
given to radiation and chemotherapy cancer patients in China to
been summarized previously Of partic-
mitigate the adverse effects of cancer therapy on the numbers
ular interest was a cluster of chemokine and cytokine genes, e.g.,
of circulating blood cells. One of the major ingredients in SI–
CXCL1, CXCL2, IL6, IL11, IL8, IL24 and TGFβ2, which showed in-
Wu–Tang is fructose, as reviewed
creased expression after irradiation with 0.1 Gy of 1-GeV/n 56Fe
We have performed studies in mice to determine whether these
ions in the absence, but not in the presence, of 5 μM SeM in the
compounds in traditional Chinese medicine were able to mitigate
medium before and during the radiation exposure
reduced circulating blood cell counts following γ -ray or SPE-like
It is also noteworthy that SeM has been shown to reduce
proton radiation. The main conclusions of the studies were as fol-
space radiation induced effects by mitigating stress-related signal-
lows: 1) SWT and fructose were both capable of increasing the
ing pathways and downregulating certain genes associated with
number of circulating lymphocytes in γ -ray or SPE proton irra-
cell adhesion These results suggest that
diated mice, and 2) fructose was more effective than SWT in in-
SeM is potentially useful as a countermeasure to prevent some of
creasing the number of circulating lymphocytes in γ -ray or proton
the acute inflammatory/immune responses induced by low-dose
irradiated mice These results are
HZE particle radiation.
important since there are essentially no countermeasures for lym-phocyte loss following radiation exposure. Lymphocytes are among
5.7. Granulocyte colony-stimulating factors as countermeasures
the most sensitive cells in the body to radiation exposure , and a reduced
In addition to the antioxidants, two forms of granulocyte
lymphocyte count in the circulation at post-irradiation times is ex-
colony-stimulating factors (G-CSFs), filgrastim and pegfilgrastim,
pected to result in greater susceptibility to infections. The fact that
were evaluated as countermeasures using the neutrophil count in
both SWT and fructose can be given orally makes them attractive
ICR mice irradiated with γ -rays or SPE-like protons as the biolog-
for use during space travel.
ical endpoint. The results demonstrated that exposure to SPE-likeproton radiation or γ -ray radiation at doses up to 2 Gy signifi-
5.9. Antibiotics as countermeasures for bacterial toxicity in mice
cantly decreased circulating neutrophil counts in a dose and time
exposed to SPE radiation and HS
dependent manner, which was prevented by treatment with ei-ther form of G-CSF evaluated in the study
Enrofloxacin, an antibiotic for veterinary use, was studied for its
These results indicate that both forms of G-CSFs could be
effectiveness as a countermeasure in experiments using the bacte-
a potential countermeasure for the reduced number of neutrophils
rial challenge model in mice. Enrofloxacin is available in an oral
in irradiated animals, although pegfilgrastim appears to be supe-
form but the mice in these experiments were treated with sub-
rior since its stimulatory effect on the neutrophil count was more
cutaneous drug. Control, irradiated (2 Gy), hindlimb suspended,
pronounced and lasted longer than that of filgrastim.
and irradiated and hindlimb suspended mice were treated with
In mouse studies using G-CSF (Neulasta) as a countermeasure
Pseudomonas aeruginosa bacteria at a dose that can be cleared by
for bacterial challenge toxicity in animals exposed to SPE radiation
untreated animals or, in later experiments, with a dose of bacteria
along with HS, Neulasta treatment was shown to reduce morbid-
that leads to morbidity in approximately 50% of untreated mice.
ity from 80–90% to 20–30% in γ -ray or proton irradiated C3H/HeN
With both challenge doses, the institution of enrofloxacin at the
mice exposed to HS and challenged with Pseudomonas aeruginosa
time of bacterial challenge reduced morbidity from 80–100% to
bacteria (Drew Weissman and colleagues, Unpublished data). In an
0%. This demonstrated the complete effectiveness of prophylactic
A.R. Kennedy / Life Sciences in Space Research 1 (2014) 10–43
antibiotic treatment in the protection of irradiated and hindlimb
many of the pigs with this condition developed non-
suspended mice against bacterial toxicity with the potential to lead
productive coughs. Thoracic radiographs and diagnostic CT scans
to morbidity/death (Drew Weissman, Unpublished data).
were performed, and the CT findings were consistent with an acute
While the effectiveness of the antibiotic countermeasure cannot
lung injury concurrent with chronic bronchial changes and vol-
be surpassed, it does carry a number of potential adverse effects,
ume loss. Differential diagnoses for these findings were radiation
especially, if it will need to be used multiple times, including the
induced lung injury or atypical infectious bronchopneumonia. To
generation of antibiotic resistant bacteria. Thus, it is believed that
help differentiate between these possibilities, sequential antibiotic
alternative countermeasures for bacterial toxicity, with different
therapy, which included atypical (e.g., mycoplasma), Gram-positive
mechanisms of action than those of antibiotics, will be useful for
and Gram-negative coverage, was initiated. After two weeks of
space travel in future exploration class missions.
therapy, it was concluded that antibiotics were not effective treat-ments for the condition. Corticosteroid therapy then began and the
5.10. Countermeasures for radiation induced emesis in ferrets
pig symptoms improved rapidly, with dramatic improvement andresolution of imaging abnormalities observed within one month.
In studies performed with ferrets, 5-HT3 receptor antagonists,
It was concluded that corticosteroids were effective treatments for
such as Zofran, have been shown to prevent or ameliorate vom-
the pneumonopathy and/or pneumonitis that developed in the pigs
iting and retching in ferrets following proton radiation exposure
exposed to SPE-like radiation
As Zofran is maintained on the ISS for nauseaand vomiting, it is the recommended 5-HT3 receptor antagonist to
5.13. Mometasone as a countermeasure for SPE proton radiation
prevent or mitigate radiation induced nausea and vomiting during
induced skin lesions in pigs
space travel.
Pigs were exposed to either a 5 Gy or 10 Gy dose of SPE-like
5.11. Countermeasures for altered bleeding times in ferrets after SPE
electron irradiation, and seven types of creams were applied to
the trunk of the animal in 1 inch2 patches covered with Tega-derm dressing consecutively for 14 days immediately after the
It is well established that endotoxin (lipid A portion) released
radiation exposure. The only cream that appeared to mitigate
by Gram negative bacteria activates certain factors of the intrin-
the radiation-induced initial hyperpigmentation was the cream
sic coagulation cascade, such as Factor XII, which in turn initiates
that contained corticosteroids (mometasone cream [Elecon]), as
fibrin formation and increases PT/aPTT values, thereby increasing
compared to the other creams, which were water-based emol-
the risk of DIC development. To counteract the factor deficiencies,
lients, a platelet growth factor containing cream, a triple antibi-
BeneFIX (recombinant Factor IX) was evaluated as a countermea-
otic containing cream, or the untreated area that was not ex-
sure for the proton radiation induced activation of the coagulation
posed to any cream other than the Tegaderm dressing. At 14 days
cascade. Treatment with BeneFIX reduced the clotting times in ir-
post-irradiation, biopsies were performed that revealed decreased
radiated ferrets back to levels that were essentially equivalent to
melanosomes, necrotic keratinocytes and melanin deposition in the
those of the non-irradiated control ferrets
areas of irradiated skin treated with mometasone compared to
Since treatment with BeneFIX increases the concentration of Factor
the untreated irradiated skin. Thus, it was concluded from these
IX, a factor depleted post-irradiation, BeneFIX could have beneficial
studies that topical application of steroids mitigate skin toxicity
effects on coagulation when administered after the radiation expo-
produced by exposure to SPE-like proton radiation (Cengel, K.A. and
Sanzari, J.K ., Unpublished data).
Phytonadione (vitamin K) is essential for post-translational
modification of a glutamate to a carboxylated-glutamate that isnecessary for Factor II, VII, IX, and X It
5.14. Transparent film dressing for protection against proton therapy
was also evaluated as a countermeasure for activation of the pro-
induced skin reactions in humans
ton radiation induced coagulation cascade. The results indicatethat phytonadione had a minor beneficial effect on PT values in
Patients undergoing proton therapy for prostate cancer fre-
the proton irradiated ferrets, but did not affect the aPTT values
quently develop radiation dermatitis. It has been reported that two
prostate cancer patients undergoing proton cancer therapy at the
Treatment of DIC with blood clotting factors or other compo-
University of Pennsylvania had radiation dermatitis that appears
nents of plasma is a topic that has been reviewed previously
to have been substantially diminished by the presence of trans-
We have demonstrated that SPE-like proton radia-
parent film dressings (Beekley stickers) In
tion led to hypocoagulability by activating the clotting cascade that
these studies, small circular (2.5 cm diameter) transparent adhe-
consumes factors involved in coagulation. SPE-like proton radia-
sive markers were placed on their skin to assist with daily align-
tion can also cause leucopenia and severe lymphocytopenia, which,
ment in these patients treated with a total proton dose of 79.2 Gy
in combination with the effects of the SPE-proton radiation on
in 1.8 Gy fractions, using two opposed lateral beams daily. It was
hemostasis, could have major health consequences in irradiated
observed that the covered areas of the skin exhibited considerably
subjects. Our studies have shown that BeneFIX can serve as a po-
diminished radiation dermatitis compared to the uncovered areas
tential countermeasure for the increased bleeding times in ferrets
of the skin, and this difference persisted for at least one month
caused by exposure to SPE-like proton radiation.
after the therapy period ended. A phantom dosimetric study wasperformed to evaluate the impact of the transparent film dressing
5.12. Corticosteroid as a countermeasure for radiation induced
on a beam's SOBP, and the results indicated no gross dosimetric ef-
pneumonitis and pneumonopathy in pigs
fect. Thus, the transparent adhesive markers appear to have atten-uated radiation dermatitis in these two patients without affecting
Several pigs exposed to simulated SPE radiation developed
the SOBP. It is hoped that this finding can improve proton-related
symptomatic, radiation-associated pneumonopathy that radio-
radiation dermatitis in other types of treatments for cancer as well
graphically involved all lung fields but was worse in the pleura and
(i.e., in other conditions in which proton radiation exposure could
apices where the radiation dose was found to be highest
lead to radiation dermatitis).
A.R. Kennedy / Life Sciences in Space Research 1 (2014) 10–43
in ferrets exposed to SPE protons and γ -radiation and on peripheral hematopoietic cell (WBC, lymphocyte
In recent years, we have been engaged in in vitro and ani-
and neutrophil) counts in minipigs exposed to simulated proton
mal studies on acute biological effects of the types of radiation at
SPE or simulated electron SPE radiation (6 + 12 MeV electrons),
the energies, doses and dose-rates relevant to space travel. Three
which resulted in a comparable dose distribution to the proton
species of animals, i.e., ferrets, pigs and mice, were used in our
SPE radiation expected in an SPE, especially at the low end of
studies to evaluate the effects of radiation on various biological
the radiation dose range evaluated The re-
endpoints including survival, cancer development, hematopoietic
sults of other studies described here suggest that the effects of
cell counts, emesis, blood coagulation, CNS endpoints (which in-
proton radiation were comparable to those of the reference ra-
cluded social exploration, submaximal exercise treadmill, and lo-
diations evaluated. These studies include studies on the lack of
comotor activity), cataract development, oxidative stress, gastroin-
bacterial containment in the GI tract from exposure to γ -rays
testinal tract bacterial translocation, immune activation, and gene
or SPE proton radiation in mice on fatigue in
expression associated with programmed cell death and ECM re-
mice after irradiation with γ -rays or protons
on the skin changes resulting from exposure to SPE
For 30-day survival, 1-GeV/n proton radiation appeared to be
proton or electron radiation on the develop-
more lethal than X-rays to mice but
ment of DIC in irradiated ferrets and on
SPE-like proton radiation was comparable to γ -rays for ferrets
mortality of mice exposed to radiation alone or radiation with
Ferrets are considerably more sensitive than
exposure to microgravity, along with a bacterial challenge (Drew
mice to the lethal effects of radiation exposure. In experiments
Weissman and colleagues, Unpublished data). Overall, the differ-
with mice irradiated at lower doses of protons (3 Gy) or HZE parti-
ences in the effects of the proton and reference radiation were
cles (0.5 Gy), acute effects of the radiation doses were not observed
relatively small or limited to the lower end of the dose range
over a 30 day experimental period, but the long-term survival in
(∼ 0.5 Gy) evaluated.
the irradiated mice was reduced significantly by the radiation ex-
The mechanism(s) for mortality in irradiated mammals is not
posure, and the decreased long-term survival was accompanied by
well understood, although numerous hypotheses have been pro-
a significant increase in the rate of development of malignant lym-
posed. At the lowest total body radiation doses leading to mor-
phomas and Harderian gland tumors, as well as the fractions of
tality, death occurs from the hematopoietic syndrome, which is
animals with malignant lymphoma or rare tumors
believed to result from the cell killing effects of radiation in the
bone marrow that lead to low numbers of circulating blood cells
With respect to peripheral WBC and lymphocyte counts, the ef-
and the resultant hematopoietic symptoms, such as infection and
fects of proton irradiation given as a homogeneous dose to the
bleeding due to the loss of leukocytes and platelets. Over the last
mice is not affected by the dose fractionation, dose-rates or proton
half century, the radiation dose required to kill half of the irradi-
energy in the ranges evaluated. In contrast, simulated hypogravity
ated subjects, known as the LD50, has been used as a parameter
brought about by PWS was shown to potentiate splenic lympho-
of radiation sensitivity for comparisons among various mammalian
cytes to the cell killing effects of radiation
species. It is well-known that the LD50 value is highly variable
A number of other studies indicated that SPE radiation and sim-
for different mammalian species; however, the bone marrow cell
ulated microgravity produced by HS led to synergistic adverse ef-
sensitivity to ionizing radiation is remarkably similar among dif-
fects on hematopoietic and immune cell functions, including bacte-
ferent species, strains and individuals
rial containment in the GI tract T-cell activation
These results suggest that the lethal ef-
and death from a bacterial challenge
fects of radiation on bone marrow and hematopoietic cells may not
at a sub-lethal dose
be the primary mechanism for radiation exposure related death.
For emesis in ferrets, the risk of SPE radiation-induced vom-
Our studies in ferrets irradiated with SPE-like protons have sug-
iting is low and may reach statistical significance only when the
gested that radiation induced activation of the coagulation cascade
radiation dose reaches 1 Gy or higher. The ED10 and ED50 values
may result in DIC, which could be a major mechanism by which
estimated for the fraction of animals that retched or vomited af-
relatively low doses of radiation lead to mortality
ter proton irradiation at the high dose rate (0.5 Gy/minute) were
The human LD50 values for radiation induced death
lower than the lower limits of the respective 95% confidence inter-
are imprecise because there have been relatively few cases in
vals established for γ -rays, suggesting that high dose rate proton
which human subjects were exposed to radiation at doses near
irradiation was more effective than high dose rate γ -ray irradia-
the LD50, especially in the last half century. The estimated human
tion in inducing retching and vomiting. There was a large sparing
LD50 value for ionizing radiation ranges from 3 to 4 Gy for young
effect observed at the low dose rates expected for SPE radiation,
adults, without medical intervention to 2 to 3 Gy for the very
such that the results for vomiting and retching in response to low
young or the old Remarkably different
dose-rate exposure to SPE radiation were not statistically signif-
LD50 values have been reported for different species
icant when compared to control animals. The trend, however, in
the experiments performed at the low dose rate appeared to indi-
as the most sensitive mammalian species
cate more retching and vomiting episodes in ferrets irradiated with
closely followed by dogs and pigs
protons than in the animals exposed to γ -rays at the same dose
The LD50 in Gottingen pigs is as low as 1.8 Gy and
widespread hemorrhaging is observed at death of the irradiated
The results from some of the studies described here indicated
Gottingen pigs with some evidence of DIC
that proton radiation had considerably more severe adverse effects
at doses near the LD50, such as the rapid onset of systemic in-
compared to those produced by comparable doses of the reference
flammation (C-reactive protein, fibrinogen) and multi-organ dys-
radiations used, which include x-rays, γ -rays and electrons. These
function Similarly, dogs exhibit hemor-
include studies on mouse survival after irradiation with protons
rhagic diathesis at doses near the LD50, and die with signs re-
and x-rays on ferret hematopoietic
sembling DIC While DIC
cell counts after exposure to SPE protons and γ -rays
has not been diagnosed as a cause of radiation induced death
on ferret retching and vomiting induced by SPE pro-
in the pig or dog studies or in irradiated human populations, a
tons and γ -rays on blood clotting times
hallmark of DIC, i.e. hemorrhage, at death has been frequently
A.R. Kennedy / Life Sciences in Space Research 1 (2014) 10–43
observed in irradiated mammals, including humans. There is ex-
for the mammalian species. This hypothesis provides a novel and
tensive evidence that widespread hemorrhages occurred in the
reasonable explanation for the great variability observed in LD50
Hiroshima and Nagasaki atomic bomb casualties, even in the rel-
values among different mammalian species.
atively low radiation dose groups with the
There is a considerable amount of evidence that humans ex-
estimated LD50 values of approximately 2.5
hibit hemorrhaging at doses near the LD50. The LD50 of the atom
Other information about hemorrhaging in hu-
bomb casualties has been estimated to be approximately 2.5 Gy
mans after irradiation comes from accidental exposures in Norway
The percent of those dying
from exposure to the atom bombs with evidence of hemorrhage
in which several people were accidentally exposed
can be estimated from the atom bomb casualty data, which has
to whole body irradiation at doses near the human LD50 and
been reviewed by The casualty dose esti-
widespread hemorrhages were observed.
mates are given in a report by the U.S. Atomic Energy Commission
A reduction in the number of platelets can result in hemorrhag-
As described by people ex-
ing and death; however, our studies suggest that the blood clotting
posed to radiation from the atom bomb used in Hiroshima have
abnormalities in irradiated ferrets is not caused by a reduction
been classified into several groups. In Group II (patients dying dur-
in the number of platelets
ing the third, fourth, fifth and sixth weeks or surviving with severe
A similar phenomenon has been reported
clinical symptoms), some of the people lived and some of the peo-
in irradiated dogs in which the platelet counts are not depressed
ple died. It is expected that the people who died in Group II were
in dogs at a "preterminal" stage in which bleeding abnormalities
those who received a dose of radiation near the human LD50 level.
are observed, although the platelet counts are decreased greatly
The data that were collected from each person represented one tis-
in irradiated dogs with full-blown DIC In
sue section from each tissue/organ examined, and the fraction of
atom bomb casualties, widespread hemorrhages were observed in
tissues exhibiting hemorrhage is reported by
people before the level of platelet counts had fallen to levels ex-
For many of the organs/tissues (e.g., kidney, liver and other or-
pected to cause hemorrhage Furthermore,
gans), the percentages exhibiting hemorrhage are as high as 60%,
while platelet injections and other blood clotting factors can have
but it is not clear what fraction of exposed individuals experienced
beneficial effects on hemorrhaging, platelet infusions do not pre-
hemorrhaging in one or more organs. (For example, it is not indi-
vent all deaths from the radiation exposure
cated whether 60% of the livers exhibiting hemorrhage are from
the same people as 60% of the kidneys exhibiting hemorrhage).
Hemorrhaging and signs of DIC have been frequently reported
Therefore, it is assumed that 60% of the people exhibiting hemor-
in higher mammalian species, such as dogs and pigs; however,
rhage is a low estimate and that the true value lies between 60%
hemorrhaging has not been reported in mice at doses near the
and 100% of the people dying in Hiroshima (after exposure at a
50. Mice are the animals that are most often used in radiobi-
50) that had evidence of hemorrhage. These data
ology studies. Bacteremia is the leading cause of death in mice
indicate that the atom bomb casualties are likely to have had very
high frequencies of hemorrhaging at death, similar to those ob-
orrhage is thought to be the major cause of death in dogs, rab-
served for other large mammals (e.g., pigs and dogs).
bits, guinea pigs or pigs
It is also noteworthy that knowledge obtained about the ad-
after irradiation at doses near the LD
verse health effects of inhomogeneous doses of interventional and
species. The pathophysiology of the hematopoietic syndrome in
therapeutic radiation is relevant for evaluation of radiation ef-
pigs is thought to be similar to that observed in humans
fects in people resulting from exposure to interventional radiology
Such differences suggest that we need new ways
procedures or therapeutic radiation, in which the skin doses are
of thinking about mechanisms for death after irradiation at doses
higher than the internal organ doses. An example of the rele-
vance for patients on earth is that high doses of SPE-like radiation
50 level. Since the human LD50 is closer to the LD50 of
ferrets, dogs and pigs than to the LD
primarily to the skin with minimal to no significant doses to in-
50 of mice, species such as fer-
rets, dogs and pigs are likely to be better animal models than mice
ternal organs have been shown to produce adverse health effects
for evaluating the effects of radiation and radiation countermea-
in internal organs (e.g., pulmonary toxicities or pneumonitis withcoughing in the lungs, bone marrow changes and reductions in cir-
sures on bleeding, coagulation cascade activation and DIC risks for
culating blood cell counts
It is noteworthy that the LD50 values for these larger mammals
7. Summary concerning the major effects of SPE and space
are considerably lower than those observed in various strains of
mice Examples of some published LD50values for mammals are as follows: Ferrets – 1.5 Gy
7.1. Dosimetry
Gottingen pigs – 1.8 Gy pigs – av-erage value – 2.6 Gy dogs – average value
With the incorporation of modern radiation oncology ap-
– 2.6 Gy monkeys – 5.07 Gy
proaches, such as CT based Monte Carlo dosimetry, into the Human
mice – average value – 8.16 Gy
Space Program, it has become feasible to accurately predict organ
Using published data, we have compared the per-
specific radiation doses for astronauts exposed to SPE radiation.
centage of animals exhibiting hemorrhage at death (from exposure
Depending on the organ system of interest (deep vs. superficial)
to radiation doses near the LD50 level) as a function of the LD50
and the fluence/energy profile of the exposure (hard vs. soft event),
of the species and observed an excellent correlation between the
either the physical size of the astronaut or the fluence/energy pro-
percent of animals developing "DIC" (from hemorrhaging at death,
file for the SPE can be the determining factor for radiation induced
the hallmark of DIC) and the LD50 value for the species/strain
(Krigsfeld, G.S. and Kennedy, A.R., Unpublished data). These fig-ures for animal hemorrhaging at death range from 100% of ferrets
7.2. Adverse effects in hematopoietic/immune system cells
– to 0% of the mice (as mice irradiated at radiation doses neartheir LD50 levels die from bacteremia
Significant decreases in WBC counts were observed in mice and
Based on this analysis, it is hypothesized that
ferrets irradiated with SPE-like proton and γ -ray radiation at to-
the propensity to develop DIC has a role in determining the LD50
tal body doses of 0.5 to 2 Gy, and in pigs irradiated with proton
A.R. Kennedy / Life Sciences in Space Research 1 (2014) 10–43
simulated SPE radiation at skin doses of 2 to 10 Gy and elec-
7.6. Disseminated intravascular coagulation
tron simulated SPE radiation at skin doses of 2.5 to 25 Gy. At thehigher doses of proton or γ -ray radiation, the neutrophil counts in
Both SPE-like proton and γ -ray radiation at doses near 2 Gy re-
the blood of both mice and ferrets, but not pigs, reached critically
sulted in DIC in ferrets, which led to 100% mortality. The threshold
low levels that, if observed in a patient in a hospital (e.g., follow-
dose of radiation for this effect is 1.5 Gy. This is an important find-
ing radiation or chemotherapy for cancer), would trigger a medical
ing, as DIC has not been established previously as a cause of death
response and suggest the use of countermeasures to increase the
in mammals following radiation exposure. The blood clotting ab-
level of neutrophils. In the pigs exposed to proton simulated SPE
normalities in ferrets were observed at a very low dose (0.25 Gy,
radiation, the neutrophil counts did not return to normal levels
which was the lowest dose evaluated in the studies), and radia-
even at months after the radiation exposure.
tion at the SPE-like low dose rate resulted in more severe effectson the blood clotting parameters in ferrets than the high dose-rate
7.3. Adverse effects on the immune system
Yucatan minipigs exposed to a 2.5 Gy dose of radiation were
also diagnosed with DIC and died (or were euthanized). Minipigs
At a dose of 2 Gy, SPE-like proton radiation or γ -ray radiation
exposed to a 2 Gy dose of SPE like proton radiation did not die,
was shown to cause breaks in the epithelial layer of the small in-
but they exhibited severe clotting abnormalities (increased bleed-
testine of mice, which allows translocation of bacteria and bacterial
ing times) that could be problematic during space travel.
products, such as LPS. The threshold for the morbidity/mortality
Based on the work in ferrets and pigs performed as part of the
effect in bacterial challenge studies was between 1.0 and 1.5 Gy.
space radiation studies described here, it is hypothesized that DIC
The proton radiation works synergistically with HS to result in an
may be a major cause of death in humans following exposure to
accumulation of a relatively large amount of LPS in the intervil-
relatively low doses of radiation.
lous regions of the ileum. Due to the synergistic effect from HS,the threshold doses for immune system parameters determined
7.7. Changes potentially related to the development of vision
without the HS treatment may underestimate the actual immune
system risks for astronauts during space travel involving micro-gravity conditions.
It was observed that pigs exposed to 2.5–7.5 Gy (skin dose)
Another major adverse effect observed in immune system
of simulated SPE – electron radiation (October 1972 event) exhib-
cells is the lack of, or reduction in the level of, activation in T-
ited increased opening pressure values, which lasted up to 90 days
lymphocytes in mice exposed to 1–2 Gy of SPE-like proton or
post-radiation. Other endpoints have also been evaluated which
γ -ray radiation, either with or without simulated microgravity (us-
could have significance for the findings that many astronauts ex-
ing the PWS and HS systems).
posed to long-duration space flight have exhibited vision alter-ations. As one example, increased nerve sheath diameters (docu-
7.4. Emesis (vomiting and retching)
mented by ocular ultrasound technology) have been observed inpigs exposed to SPE-like radiation. Increased nerve sheath diame-
In female descented Fitch ferrets, irradiation with 60Co γ -rays
ters have also been documented by ocular ultrasound technology
or 155-MeV protons at a high dose rate of 0.5 Gy/minute resulted
in astronauts and this finding has been implicated in the develop-
in dose-dependent changes in the endpoints that are indicative of
ment of astronaut vision alterations
retching and vomiting. The minimum radiation doses required to
A relatively low dose of SPE-like radiation (skin doses as low
induce statistically significant changes in retching- and vomiting-
as 2.5 Gy) can lead to increased intracranial pressure in pigs, and
related endpoints were 0.75 and 1.0 Gy, respectively. The RBE of
a dose of 5 Gy of proton simulated SPE radiation was shown to
the proton radiation at the high dose rate (relative to the γ -rays at
increase the nerve sheath diameter. The statistically significant ef-
the same dose rate) did not differ significantly from 1. Similar, but
fects observed on these endpoints in the pig studies suggest that
smaller and less consistent, changes in the retching- and vomiting-
astronaut exposure to these relatively low doses of SPE radiation
related endpoints were also observed for ferrets irradiated with
could exacerbate the vision alterations known to exist in astro-
γ -rays and protons at the low dose rate of 0.5 Gy/hour. Since this
nauts during future exploration class missions.
low dose rate is similar to a radiation dose rate expected duringa SPE, these results suggest that the risk of SPE radiation-induced
7.8. Threshold radiation doses for statistically significant adverse
vomiting is low and is likely to reach statistical significance only
biological effects from SPE-like radiation in vivo
when the radiation dose reaches 1 Gy or higher.
The estimated threshold radiation doses to cause significant ad-
verse biological effects in vivo for various endpoints measured are
7.5. Skin effects
summarized as follows:
At skin doses of 7.5 Gy and higher, there is a persistent im-
• Reductions in WBC counts in mice and ferrets: a homogeneous
munological dysfunction in pig skin, which is characterized by an
dose of 0.5 Gy; in pigs: a homogeneous dose of 2.0 Gy (lowest
enhanced DTH response; a similar effect was observed in mice
dose evaluated) of SPE-like proton radiation.
at doses of 2 Gy or less. At higher doses of radiation, the pig
• Levels of death in mice exposed to SPE like protons, along with
skin becomes more sensitive to touch (lymphedematous), and
HS, in response to a bacterial challenge: 1.0 to 1.5 Gy.
there can be blistering, burns and epithelial dysfunction (e.g., pig-
• Elevated levels of LPS and LPB in mouse serum: 2 Gy of SPE-
ment incontinence, which indicates defective cell to cell transfer
like proton radiation.
of biomolecules). The threshold radiation doses for the skin effects
• Increases in blood clotting time in ferrets: 0.25 Gy of SPE-like
are 4.5 to 5 Gy. The decrease in vascular bed area is the most se-
protons at a low, SPE-like dose rate.
rious skin effect potentially resulting from irradiation at very high
• Lack of T cell activation in mice: 1 Gy.
skin doses. However, this complication would be a highly unlikely
• Skin effects: skin hyperpigmentation, epidermal thickening,
occurrence for astronauts since such high radiation doses are not
and decrease in vascular bed area: 4.5 to 5 Gy of pSPE or eSPE
expected during most SPEs.
A.R. Kennedy / Life Sciences in Space Research 1 (2014) 10–43
• Emesis in ferrets (for high dose-rate irradiation): 1 and 0.75 Gy
7.10. Dose-rate effects
for vomiting and retching, respectively. For low SPE like dose-rates, the results were not statistically significant at doses up
One major endpoint evaluated that was affected by dose rate
to 2 Gy for SPE like proton radiation compared to controls.
was ferret emesis, which was significantly increased after irradia-
• Levels of death from, or signs of, DIC in ferrets or pigs. All fer-
tion with SPE-like proton radiation or γ -ray radiation administered
rets died with signs of DIC at a dose of 2 Gy from SPE-like
at the high dose rate of 0.5 Gy/minute, but not at a low dose rate
protons or γ -ray radiation. The threshold dose for this effect
of 0.5 Gy/hour. The other major endpoint affected by dose-rate
is 1.5 Gy. Ferrets are thought to be more susceptible to radi-
was the blood clotting time, which was increased to a considerably
ation induced DIC than humans, while the sensitivity of pigs
greater extent in the studies with the low, SPE-like dose-rate than
to radiation induced DIC may be more like that of humans. In
in the high dose-rate radiation studies. For all other endpoints, the
an experiment in which 3 Yucatan minipigs were exposed to
sparing effects of the radiation dose rate are either insignificant or
X-rays at a dose of 2.5 Gy, one pig died and another pig was
minimal and not biologically meaningful.
euthanized; both of these pigs had evidence of DIC. 2.5 Gy is
8. Agents identified as countermeasures for space radiation
a potentially lethal dose of radiation for humans as well. At
induced adverse biological effects
an SPE-like radiation dose of 2.0 Gy, pigs had severe clottingabnormalities suggestive of DIC. With the other space stres-
As a part of the work for this program, the following agents
sors (e.g., microgravity, elevated levels of oxygen during EVAs
have been identified as countermeasures, which may lead to risk
and carbon dioxide in the spacecraft, etc.), the lethal human
reductions in astronauts exposed to space radiations.
dose could be < 2.5 Gy and astronauts may be susceptible to
A. Dietary antioxidants fructose
the onset and progression of DIC at doses that could be re-
and G-CSFs (Neupogen, Neulasta)
ceived from exposure to SPE radiation. In any case, astronauts
as countermeasures to prevent or alleviate the loss of circulating
are likely to have bleeding abnormalities from very low SPE
white blood cells
radiation doses (25 cGy or below), and they should avoid ex-
posure to anything that could damage the skin integrity due
to the bleeding risk following a sizeable exposure to SPE radi-
B. Orally administered antibiotics (e.g., enrofloxacin) as a coun-
termeasure to prevent or alleviate translocation of bacteria andbacterial products as well as death from bacterial challenge in ani-
7.9. RBE values
mals exposed to SPE-like proton radiation and hindlimb suspension(Drew Weissman, Unpublished data).
A higher RBE value for a given effect indicates that a more se-
C. 5-HT3 antagonists (e.g., oral ondansetron [Zofran]) as a coun-
vere effect is expected for exposure to SPE radiation than from
termeasure for proton radiation induced emesis (retching and
conventional reference radiations (e.g., γ -rays, x-rays or electrons).
In our studies, it was observed that there can be: 1) different RBEs
D. Corticosteroid therapy as a countermeasure for pneumonopa-
for different biological endpoints in the same animal species/strain,
thy/pneumonitis that developed after radiation exposure
and 2) different RBEs for the same biological endpoint in differ-
E. Topically applied steroid cream (mometasone-Elecon) (Cen-
ent species/strains. However, for most of the endpoints evaluated,
gel, K.A. and Sanzari, J.K., Unpublished data) and transparent film
which include hematopoietic blood cell counts, immune system
dressing as countermeasures for radiation in-
parameters and fatigue measured in mice, emesis evaluated in
duced skin damage (e.g. hyperpigmentation).
ferrets, skin effects assessed in pigs, the RBE values were not sig-
F. Dietary antioxidants as a countermeasure to decrease the risk
nificantly different from 1 except for the following:
of long-term radiation effects, e.g. cancer
Hematopoietic blood cell counts in ferrets: RBE ranges from
G. Benefix as a countermeasure for increased bleeding times
1.2 to 1.6 for the WBC count at 48 hours after irradiation, RBE
after radiation exposure
ranges from 1.9 to 2.1 for neutrophil count at 48 hours afterirradiation (RBE values were particularly increased at the low
end of the SPE proton dose range evaluated);
• Hematopoietic blood cell counts in pigs: RBE ranges from 2.4
Most of the research investigations from the author's labo-
to 4.1 for the WBC count, and 2.2 to 5.0 for the neutrophil
ratory discussed in this review were supported by the National
count on day-4 post-irradiation (RBE values were particularly
Space Biomedical Research Institute (NSBRI). The NSBRI is funded
increased at the low end of the SPE proton dose range evalu-
through NASA NCC 9-58. In addition, some studies were funded by
a NASA grant (NAG9-1517). Several trainees participating in theseinvestigations were supported by the NIH-funded Radiation Biol-
RBE values were determined for hematopoietic cell counts in
ogy Training Grant (T32CA009677).
mice, ferrets and pigs; therefore, they can be compared acrossthese three species. From these data, it was observed that the RBE
values of the SPE-like radiation for white blood cell counts varygreatly between mice, ferrets and pigs, with the RBE values being
somewhat greater in ferrets than in mice, and considerably greater
in pigs than in ferrets or mice. This trend suggests that the RBE
values of SPE radiation for white blood cell counts could be con-
siderably greater in humans than those observed in smaller mam-
mals, and that SPE proton radiation may be far more hazardous
to humans than previously estimated from studies performed in
small animals (e.g., rodents).
A.R. Kennedy / Life Sciences in Space Research 1 (2014) 10–43
Czarnik, T.R., 1988. Medical Emergencies in Space.
A.R. Kennedy / Life Sciences in Space Research 1 (2014) 10–43
Hlatky, L., Hahnfeldt, P., 2014. Beyond the cancer cell: progression-level determi-
nants highlight the multiscale nature of carcinogenesis risk. Cancer Res. 74 (3),
659–664. Epub 2013 Nov 22.
PMID: 24272486.
A.R. Kennedy / Life Sciences in Space Research 1 (2014) 10–43
Krigsfeld, G.S., Savage, A.R., Billings, P.C., Lin, L., Kennedy, A.R., 2014. Evi-
dence for radiation induced disseminated intravascular coagulation as a major
cause of radiation induced death in ferrets. Int. J. Radiat. Oncol. Biol. Phys.
pii S0360-3016(13)03611-0 [Epub
ahead of print].
A.R. Kennedy / Life Sciences in Space Research 1 (2014) 10–43
NCRP, 1989. National Council on Radiation Protection and Measurements (NCRP)
Report No. 98. Guidance on radiation received in space activities. NCRP.
NCRP, 2006. National Council on Radiation Protection and Measurements (NCRP)
Report No. 153. Information needed to make radiation protection recommenda-
tions for space missions beyond low-earth orbit. NCRP.
NCRP, 2010. National Council on Radiation Protection and Measurements (NCRP)
Report No. 168. Radiation dose management for fluoroscopically-guided inter-
ventional medical procedures. NCRP.
NCRP, 2011. National Council on Radiation Protection and Measurements (NCRP) Re-
port No. 170: Second primary cancers and cardiovascular disease after radiation
therapy. NCRP.
A.R. Kennedy / Life Sciences in Space Research 1 (2014) 10–43
Romero-Weaver, A.L., Ni, J., Lin, L., Kennedy, A.R., 2014. Orally administered fruc-
tose increases the numbers of peripheral lymphocytes reduced by expo-
sure of mice to gamma or SPE-like proton radiation. Life Sci. Space Res.
Sanzari, J.K., Wan, X.S., Rusek, A., Diffenderfer, E.S., Kennedy, A.R., 2014. Acute
hematological effects in mice exposed to the expected doses, dose-rates, and
energies of solar particle event-like proton radiation. Life Sci. Space Res.
A.R. Kennedy / Life Sciences in Space Research 1 (2014) 10–43
Source: http://alpha.sinp.msu.ru/~panov/Lib/Papers/MARS/1-s2.0-S2214552414000108-main.pdf
Available Online through www.ijpbs.com (or) www.ijpbsonline.com IJPBS Volume 3 Issue 4 OCT-DEC 2013 255-264 Research Article Pharmaceutical Sciences METHOD DEVELOPMENT AND VALIDATION OF RP-HPLC METHOD FOR SIMULTANEOUS ESTIMATION OF DICYCLOMINE HYDROCHLORIDE AND DICLOFENAC POTASSIUM IN TABLET DOSAGE FORMS
Diseases and ConditionsUlcerative colitis By Mayo Clinic Staff Ulcerative colitis (UL-sur-uh-tiv koe-LIE-tis) is an inflammatory bowel disease (IBD) thatcauses long-lasting inflammation and ulcers (sores) in your digestive tract. Ulcerativecolitis affects the innermost lining of your large intestine (colon) and rectum. Symptomsusually develop over time, rather than suddenly.