Angair.org.au



Author's personal copy
International Journal for Parasitology: Parasites and Wildlife 3 (2014) 88–94
Contents lists available at ScienceDirect
International Journal for Parasitology:
Parasites and Wildlife
Experimental manipulation reveals few subclinical impacts of a parasitecommunity in juvenile kangaroos
Jemma Cripps a,b,⇑, Ian Beveridge b, Richard Ploeg b,1, Graeme Coulson aa Department of Zoology, The University of Melbourne, VIC 3010, Australiab Faculty of Veterinary Science, The University of Melbourne, Veterinary Clinical Centre, Werribee, VIC 3030, Australia
Large mammalian herbivores are commonly infected with gastrointestinal helminths. In many host spe-
Received 27 January 2014
cies, these helminths cause clinical disease and may trigger conspicuous mortality events. However, they
Revised 15 March 2014
may also have subclinical impacts, reducing fitness as well as causing complex changes to host growth
Accepted 28 March 2014
patterns and body condition. Theoretically, juveniles should experience significantly greater costs fromparasites, being immunologically naive and undergoing a significant growth phase. The aims of our studywere to quantify the subclinical effects of helminths in juvenile eastern grey kangaroos (Macropus gigan-
teus), which commonly harbour large burdens of gastrointestinal nematodes and are susceptible to asso-
ciated mass mortality during cold, wet conditions. We conducted a field experiment on a population of
free-ranging kangaroos, removing nematodes from one group of juveniles using an anthelmintic treat-
ment. We then compared growth parameters (body condition and growth rates) and haematological
Macropus giganteus
parameters of this group with an age-matched, parasitised (untreated) control group. Treated juvenile
kangaroos had significantly higher levels of plasma protein (albumin) but, contrary to our predictions,showed negligible changes in all the other parameters measured. Our results suggest that juvenile kan-garoos are largely unaffected by their gastrointestinal helminth burdens, and may be able to compensatefor the costs of parasites.
! 2014 The Authors. Published by Elsevier Ltd. on behalf of Australian Society for Parasitology. This is an
open access article under the CC BY-NC-ND license (http://creativecommons.org/licenses/by-nc-nd/3.0/).
in livestock and wildlife often differ, due to the difficulties ofstudying natural host–parasite relationships and quantifying fit-
Gastrointestinal helminths commonly infect mammalian herbi-
ness consequences. In order to thoroughly investigate such effects,
vores (Sykes, 1987). Helminth infections often have clinical
experimental manipulation is imperative. Field experimentation
impacts, causing disease and mortality (Holmes, 1987), but can
allows the actual costs of parasites on hosts to be investigated rig-
also cause what has been termed ‘subclinical' disease (Gunn and
orously, eliminating many of the issues associated with extrapolat-
Irvine, 2003), inducing more subtle effects in the host. Subclinical
ing laboratory results onto free-living individuals or populations
impacts are well known in livestock: reductions in appetite and
(Seitz and Ratte, 1991). Studies of fitness consequences in the wild
food absorption caused by helminth infections can decrease host
typically focus on natural covariation between parasite load and
fecundity and growth (Mejia et al., 1999), body condition
fitness parameters, and so may be confounded by the inherent dif-
(Loyacano et al., 2002) and alter metabolism (O'Kelly et al.,
ferences among individuals that can contribute to high parasite
1988). In contrast, the effects of parasitism on wildlife have
burdens. Consequently, it is often unclear whether changes in fit-
received far less attention, although there is mounting evidence
ness parameters are due to heavy parasite burdens, or if these bur-
that parasites can have similar negative impacts, reducing fitness
dens result from other pre-existing factors related to fitness.
(Watson, 2013) as well as causing complex changes to host physi-
Ecological host–parasite studies in wild animals have been mostly
ology (Van Houtert and Sykes, 1996), behaviour (Scantlebury et al.,
based on correlations, although there have been some field exper-
2007) and population dynamics (Hudson et al., 1992a; Albon et al.,
iments in wild systems (e.g. Svalbard reindeer, Rangifer tarandus
2002; Stien et al., 2002). However, the types of impacts measured
platyrhynchus (Stien et al., 2002), Soay sheep, Ovis aries (Gulland,1992) and red grouse Lagopus lagopus scoticus (Hudson et al.,1992b)).
⇑ Corresponding author at: Department of Zoology, The University of Melbourne,
VIC 3010, Australia. Tel.: +61 3 8344 4862.
The subclinical effects of parasites can be extremely difficult to
E-mail address: [email protected] (J. Cripps).
quantify in the wild. Ecologists tend to use body condition as an
1 Previous address: IDEXX Laboratories, Mount Waverley, VIC 3149, Australia.
http://dx.doi.org/10.1016/j.ijppaw.2014.03.0052213-2244/! 2014 The Authors. Published by Elsevier Ltd. on behalf of Australian Society for Parasitology.
This is an open access article under the CC BY-NC-ND license (http://creativecommons.org/licenses/by-nc-nd/3.0/).
Author's personal copy
J. Cripps et al. / International Journal for Parasitology: Parasites and Wildlife 3 (2014) 88–94
indicator of an animal's health and reproductive potential. Body
the pouch, young will exit permanently at around 320 days
condition essentially reflects available energy reserves (Green,
(Poole, 1975). Toward the end of pouch life, juveniles begin to for-
2001): an animal in good condition is assumed to have more
age on the pasture and are exposed to the infective stages of nem-
reserves than one in poor condition (Schulte-Hostedde et al.,
atodes. Juveniles will continue to associate with and suckle from
2005). Energy reserves can be quantified directly by measuring
their mothers until over 18 months of age (Poole, 1975). The year
fat stores (e.g. amount of back or kidney fat (Riney, 1955)) or
following permanent pouch exit is the most critical for juvenile
non-invasively using mass/size ratio indices of body condition,
kangaroos, as they must undergo substantial growth. During this
which attempt to determine the size of energy stores after correct-
period, the average monthly weight gain is 1.4 kg for males and
ing for structural body size (Schulte-Hostedde et al., 2005). Alter-
0.9 kg for females (Poole et al., 1982). To sustain this growth, juve-
natively, haematological and serum biochemical parameters can
niles have around 1.8 times the energy requirement of mature,
be used to assess an animal's health. Although less commonly used
non-lactating females (Munn and Dawson, 2004). In addition, dur-
in ecological studies of wildlife, haematological parameters may
ing this period of growth, individuals are immunologically naive
provide more sensitive information on the immediate physiologi-
(Arundel et al., 1990), and become infected by gastrointestinal par-
cal status of a host (Milner et al., 2003; Budischak et al., 2012). That
asites. Individual variability in body size increases following per-
is because parasite infections can alter haematological parameters
manent pouch exit (Poole et al., 1982), suggesting that growth
directly through haematophagy (blood-feeding) and indirectly
rate is a plastic trait that can be influenced by external factors.
through activation of host immunity in response to infection or
We examined the effect of concomitant infection with multiple
by limiting the digestion and absorption of essential nutrients,
parasites on the growth, body condition and blood chemistry of
such as protein (Colditz, 2008). Red blood cell counts, haemoglobin
one cohort of free-ranging juvenile eastern grey kangaroos, by
and plasma protein concentrations can all be used to assess an ani-
manipulating parasite loads. We removed gastrointestinal para-
mal's health and condition, and have been directly linked to perfor-
sites from a group of juveniles using an oral anthelmintic and then
mance and reproductive success (Moore and Hopkins, 2009). The
compared them with control individuals, with the expectation that
combined application of both haematological and body condition
control juveniles would show subclinical effects. We predicted that
indices may therefore provide greater insight into the subclinical
due to an increased availability of nutrients and energy resources,
effects of parasite communities on a host.
treated juveniles would have a greater growth rate and mass gain,
Juvenile mortality is commonly increased by infection (Schmidt
and would increase their body condition relative to controls. We
et al., 1979), however evidence from livestock suggests that this
also predicted that there would be changes in haematological
age-class can also experience considerable subclinical effects such
parameters, with decreases in red blood cell counts, haemoglobin
as reductions in body weight (Chiejina and Sewell, 1974), growth
concentration and haematocrit in control juveniles. Similarly, we
(Loyacano et al., 2002) and appetite (Kyriazakis et al., 1998).
expected that serum biochemistry would indicate subclinical
Despite the evidence from livestock hosts, it is unclear to what
effects, with decreased levels of total protein and albumin, and
degree subclinical effects occur in juveniles of wildlife species. The-
increases in levels of globulin.
oretically, juveniles should experience significant costs wheninfected with parasites due to the nutritional deficits they causeand the costs of mounting an immune response (Colditz, 2008),
2. Materials and methods
and these effects should be particularly marked during earlygrowth and development. Such effects are important to compre-
hend as it is well established that conditions early in life can havesignificant implications for survival and reproductive success as an
This study was conducted at the Anglesea Golf Club (38"240S,
adult (Metcalfe and Monaghan, 2001).
114"100E) in southern Victoria, Australia, in 2012. The golf course
Most wildlife hosts harbour complex parasite communities
covers an area of 73 ha and contains open, grassy fairways domi-
(Bordes and Morand, 2011), and kangaroos (Marsupialia: Macropo-
nated by couch grass (Cynodon dactylon), separated by patches of
didae) are known to support more species of parasites than any
woodland and shrubland (Inwood et al., 2008). The course is bor-
other group of mammals (Beveridge and Chilton, 2001). The east-
dered by native heathy woodland to the north and west; kangaroos
ern grey kangaroo (Macropus giganteus) carries a diverse fauna of
move freely between the course and native vegetation, as well as
gastrointestinal nematode parasites in its complex, sacculated
through residential properties in the south and east. Population
forestomach (Beveridge and Arundel, 1979), with most species
surveys (following Inwood et al., 2008) at the time of the study
showing seasonal fluctuations, peaking in the winter months
showed that the population density of kangaroos at the site was
(Arundel et al., 1990). Most of these gastrointestinal nematodes
approximately 2.0/ha (Cripps and Coulson, unpublished data).
are directly transmitted via ingestion (Sykes, 1987). Adult kanga-
Potential predators at the site include the red fox (Vulpes vulpes)
roos do not appear to develop immunity to most of these nema-
and domestic dogs (Canis lupus familiaris).
tode species (Arundel et al., 1979), and juveniles are susceptible
Post-mortem examinations of three juveniles found dead at the
to gastrointestinal parasitism, primarily from high burdens (400–
study site during 2010–2011, using standard methods (Beveridge
1500) of the intestinal trichostrongylid nematode Globocephaloides
and Arundel, 1979), revealed a diverse gastrointestinal parasite
trifidospicularis Juveniles can experience high mortality, coupled
community. The stomach contained the strongyle nematodes
with declining haematocrit and plasma protein concentrations, in
Rugopharynx macropodis (intensity 8240–69,000 per kangaroo),
their first winter post-weaning between 14 and 20 months of age
Rugopharynx rosemariae (500, n = 1) and Pharyngostrongylus kappa
(Arundel et al., 1990). Populations of eastern grey kangaroos can
(500–10,000). The small intestine contained G. trifidospicularis
reach high densities, and individuals are gregarious, forming
(31–73), levels well below those known to cause clinical impacts
mixed-sex, open-membership groups to forage and rest (Coulson,
(Arundel et al., 1990). The large intestine contained two oxyuroid
2009), conditions that favour helminth parasite transmission
nematodes, Macropoxyuris brevigularis (40–3450) and Macropoxyu-
(Altizer et al., 2003).
ris longigularis (500–1050), and one strongyle nematode species
Eastern grey kangaroos are capable of breeding throughout the
Macropostrongyloides baylisi (50–9000). The bile ducts of two juve-
year, but most births occur between September and March, during
niles had the cestode Progamotaenia festiva. Examination of seven
the austral spring/summer months (Poole, 1983). Following a short
adults from the site revealed a further ten helminth species: Labio-
gestation period and then an extended period of development in
simplex kungi, Labiosimplex bipapillosus, Cloacina pelops, Cloacina
Author's personal copy
J. Cripps et al. / International Journal for Parasitology: Parasites and Wildlife 3 (2014) 88–94
herceus, Cloacina hermes, Cloacina selene, Cloacina artemis, Cloacina
of 77 days, range 68–114 days) and final body measurements were
expansa, Cloacina obtusa, and Alocostoma clelandi in the stomach
collected. In total, each individual was captured three times.
(Cripps, unpublished data). Coinfection levels ranged from 5 to 8helminth species in juveniles kangaroos, and a further 3–5 species
2.3. Faecal egg counts
in adults (Cripps, unpublished data). Total worm counts in juve-niles (12,870–79,853) and adults (1844–84,017) were well within
To determine the efficacy of the parasite treatment, egg counts
the range recorded for eastern grey kangaroos (Arundel et al.,
were conducted on faecal samples collected within 33 days of first
1990; Beveridge and Arundel, 1979). Ectoparasites (lice, ticks)
capture. Samples were collected again at 40–90 days post-treat-
were not recorded as eastern grey kangaroos have relatively few
ment, prior to the second re-treatment period. At this point, it is
ectoparasites (Beveridge and Arundel, 1979).
possible that many of the nematodes were larval stages and thuswere not reflected in the faecal egg counts. Faecal samples were
2.2. Animal capture and treatment
collected in 2 h blocks at dawn and dusk, as this is when kangaroosare actively foraging and defaecation rates are greatest (Johnson
Juvenile kangaroos (n = 42) were first captured in March and
et al., 1987). Marked juveniles were observed until they defaecat-
April 2012. Due to their habituation to humans, kangaroos at this
ed, and observers collected faecal samples immediately after they
site tolerate close approach on foot. Juveniles were identified pri-
were deposited. It was not possible to collect samples from every
marily based on their size, but also on whether they were closely
individual, so faecal samples were collected for a subset of juve-
associated with an adult female. Juveniles were captured using
niles in each group. Samples were collected from 13 juveniles (5
either an extendable pole syringe (1.4 m, 2.4 m or 3.6 m long)
treated, 8 controls) days 12–33 post-treatment, and from 20 juve-
(King et al., 2011), or by an injection arrow fired from a band-pow-
niles (9 treated, 11 controls) days 40–90 post-treatment. Samples
ered gun (Para-medic; Wildvet, Melbourne, Victoria, Australia).
were maintained at 4 "C and analysed within 24 h. The number
Both methods injected the hind limb musculature with Zoletil#
of eggs per gram (epg) was determined by a modified McMaster
100 (100 mg/mL of 50:50 tiletamine hydrochloride – zolazepam
technique using 2 g of faeces mixed with 60 mL of saturated
hydrochloride mixture; Virbac Animal Health Pty Ltd, Milperra,
sodium nitrate solution (Redox Pty Ltd, Minto, New South Wales,
New South Wales, Australia) at a dose of approximately 5 mg/kg
Australia). An aliquot of 0.5 mL of homogenised filtrate was trans-
ferred into a Whitlock Universal counting chamber then examined
To identify individuals, they were fitted with a unique combina-
under a microscope at 100! magnification. Only typical strongylid
tion of coloured, reflective ear tags (Leader, Craigieburn, Victoria,
eggs were counted, with each egg representing 60 epg of faeces.
Australia). Standard body measurements (Poole et al., 1982) werecollected using a retractable tape measure and Vernier calipers.
2.4. Blood collection and analysis
Leg, pes (foot) and arm lengths were measured to the nearestmm; body mass was measured to the nearest 0.1 kg using 25-kg
During the second recapture, blood samples were also collected
spring scales (Salter, Melbourne, Victoria, Australia).
from the lateral caudal vein. Blood for haematology was trans-
The approximate date of birth of each individual was calculated
ferred into 2 mL vacutainers containing EDTA, and for serum anal-
from the mean of three estimates based on leg, foot and arm mea-
ysis into 5 mL vacutainers containing gel for serum separation but
surements at the first capture using growth tables provided by
no additives. Vials were immediately placed in a cooler and taken
Poole et al. (1982). In some cases, one measurement gave an esti-
back to the field base, where blood for serum analysis was centri-
mate that was >2 months apart from the other two. If this
fuged for 15 min. Blood smears were also prepared on glass slides
occurred, birthdate was calculated using the mean of the other
within 4 h of collection. Sera and whole blood samples were refrig-
two estimates. Only individuals born after 1 August 2010 were
erated until transport to a commercial, NATA-accredited, diagnos-
included in the final analysis to ensure that they were 621 months
tic veterinary laboratory (IDEXX Laboratories, Mount Waverley,
of age and therefore encountering nematode larvae for the first
Victoria, Australia). The time between blood collection and deliv-
time over the winter months of June – August 2012.
ery to the lab was <24 h. Whole blood was analysed for the number
Individuals were randomly allocated to either a control (n = 20)
of red blood cells, the haemoglobin concentration and haematocrit.
or a treatment (n = 22) group, stratified by sex to ensure equal
Assessment of haematologic values was performed using a Sysmex
numbers of males and females in each group. Treated individuals
XT-2000i haematology analyzer (Sysmex Corportaion, Kobe,
were given an oral dose of albendazole (Alben# for sheep, lambs
Japan). Serum chemistry profiles were obtained with an Olympus
and goats, 19 g/L, Virbac Animal Health Pty Ltd, Milperra, New
AU 400 analyzer (Olympus Diagnostics, Hamburg, Germany) and
South Wales, Australia) at a rate of 3.8 mg/kg body mass (Cripps
included total protein, albumin and globulin.
et al., 2013), while control individuals were left untreated. No oralcontrol was administered to untreated individuals to avoid indi-
2.5. Statistical analysis
rectly affecting the gastrointestinal fauna. A number of juvenilesdisappeared (either died or dispersed) during the study, so only
Logistic regression was used to analyse the effects of sex and
15 controls (8 male, 7 female) and 12 treatment (8 male, 4 female)
treatment on the disappearance rate of individuals throughout
kangaroos were included in the analysis. The average age of these
the study. Faecal egg count reduction calculations were made
individuals at first capture was 14.5 months (range 11–
according to Wood et al. (1995) using the Excel plug-in ‘Reso'
19.5 months).
(Cameron, 2003). Analysis of the effects of treatment on kangaroo
Juvenile kangaroos were recaptured between May and June
faecal egg counts was carried out using Genstat, Version 10 (VSN
2012 to re-administer the anthelmintic to the treated group and
International Ltd., Hemel Hempstead, UK). Faecal egg counts were
to collect body measurements from both groups. The interval of
log (1 + epg) transformed to meet the assumptions of normality
re-treatment was based on the estimated pre-patent period of
and analysed using restricted maximum-likelihood analyses
infection (approximately 3 months) in eastern grey kangaroos
(REML), with time and treatment as fixed factors, and kangaroo
(Cripps et al., 2013). Individuals were recaptured in a similar order
identity as a random factor to account for repeated measures.
to their initial captures; the average time between first and second
Differences in body mass and leg lengths of juveniles in the
capture was 77 days (range 69–91). Individuals were recaptured
treated and control groups at the initial capture were tested using
again between July and September 2012 (mean recapture interval
independent sample t-tests. The Scaled Mass Index (Peig and
Author's personal copy
J. Cripps et al. / International Journal for Parasitology: Parasites and Wildlife 3 (2014) 88–94
Green, 2009) was used to measure body condition. This index over-comes several drawbacks of other indices, many of which fail toaccount for the changing relationship between mass and lengthas growth occurs. Following Peig and Green's procedure (2009),the most strongly correlated body measurement with body mass(on a log–log scale) was determined. Initially each sex was testedseparately, but there was no difference in the strength of the cor-relations so sexes were pooled. Both leg and arm length were morehighly correlated with body mass (leg: r = 0.92, P < 0.01; arm:r = 0.91, P < 0.01) than pes length (r = 0.81, P < 0.01), so we choseleg length as the length (Li) value for each individual. The popula-tion mean (L0) was calculated separately for each of the three cap-
ture periods. The Scaled Mass Index of body condition was thencalculated as Mi(L0/Li)bSMA, where Mi is the body mass of the indi-
vidual and bSMA is the standardised major axis regression slopeof the lnMi " lnLi plot (Peig and Green, 2009).
To assess the effect of treatment across the three capture peri-
ods, repeated measure ANOVAs were performed to determine dif-ferences in body mass, leg length and the Scaled Mass Index. The
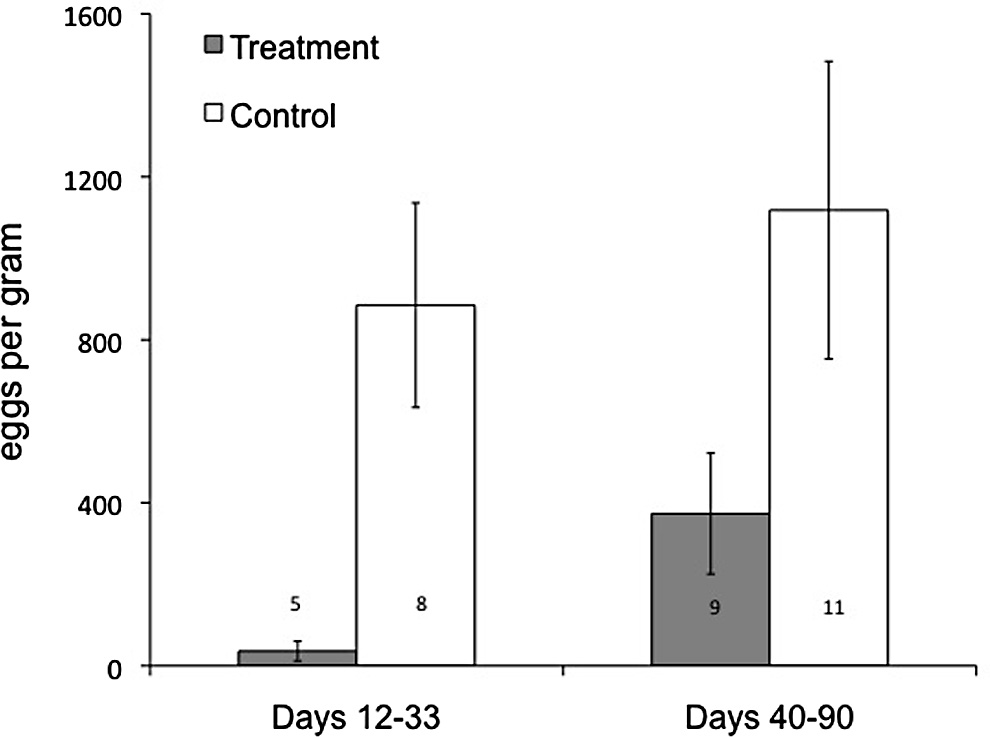
Fig. 1. Mean faecal egg counts for control and anthelmintic-treated juvenile eastern
main factor was treatment (albendazole administered or untreated
grey kangaroos in two periods post-capture (12–33 days) and initial treatment (40–90 days) at the Anglesea Golf Club, Victoria, Australia, from March to May 2012.
control), and the repeated factor was captures. Mauchly's test of
Bars indicate standard errors. It was not always possible to sample each individual
sphericity indicated that the assumption of sphericity was violated
in each period; numbers on columns indicate sample size for each time period.
in all cases (body mass: v22 = 9.86, P = 0.007; leg: v22 = 10.93,
P = 0.004; body condition: v22 = 13.97, P = 0.001), so a Greenhouse–
Geisser correction was used. Analysis of the effect of treatment on
in the faecal flotations. Eimeria oocysts were present in some sam-
haematological and serum biochemical parameters was carried out
ples but at very low numbers.
using independent sample t-tests or Mann–Whitney U tests in the
Body mass of control and treated juvenile kangaroos did not dif-
cases where data were non-normal.
fer at the time of first capture, (t25 = 0.19, P = 0.85), nor did skeletal
Power analyses were performed for each growth and blood
size, measured as leg length (t25 = 0.01, P = 0.99). Mass gain was
parameter, using G*Power (Erdfelder et al., 1996). The magnitude
explained by time (F1.49,37.39 = 21.22, P < 0.001) but not treatment
of effects for most of the parameters we tested is rarely reported
(F1,25 = 0.04, P = 0.84), and there was no interaction between the
in the literature. The magnitude of body mass effects reported for
two (F1.49,37.39 = 0.44, P = 0.59). There was also no significant effect
livestock hosts are extremely variable. When compared to parasi-
of treatment on the growth of leg length (F1,25 = 0.08, P = 0.93).
tised controls, unparasitised heifers gained approximately 18%
Instead, time explained leg growth (F1.46,36.61 = 71.39, P < 0.001),
more weight (Mejia et al., 1999), whereas in sheep, the difference
and there were no interactions between time and treatment
ranged from 74% (McLeod and Wolff, 1968) to 81% (Anderson et al.,
(F1.46,36.61 = 0.24, P = 0.72). The body condition of individual juve-
1980). Consequently, we used both 20% and 80% differences
nile kangaroos increased during the study (F1.39,34.69 = 21.60,
between the treatment and control groups as the effect sizes in
P < 0.001; Table 1) but there was no effect of treatment
these calculations.
(F1,25 = 0.01, P = 0.91) nor any interaction between treatment and
All other statistical analyses were carried out using SPSS Ver-
time (F1.39,34.69 = 0.32, P = 0.65). With the estimated 80% effect size,
sion 21 (IBM Corporation, Armonk, New York, USA). The assump-
power was high for leg measurements (>0.8, Table 1) and moderate
tions of parametric statistic analyses (normality and equality of
for body mass and body condition (0.3–0.8, Table 1). With the esti-
variances) were tested for all sets of data. Normality was assessed
mated 20% effect size, power was low (<0.3, Table 1) for all of the
using the Kolmogorov–Smirnov statistic with a > 0.05, and
Levene's test for homogeneity of variances was used to test equal-
Serum albumin levels were 8% higher in treated juveniles than
ity of variances, with a > 0.05. We did not apply sequential Bonfer-
in controls, and this difference was significant (P = 0.01, Table 2).
roni adjustments on the basis that we report power analyses for all
While there were no significant differences in the concentrations
our tests, and had already selected a subset of blood and growth
of total protein, globulin or haemoglobin, the red cell count or
parameters for analysis (Moran, 2003).
the haematocrit, all of these parameters showed trends in the pre-dicted directions, with lower levels in the parasitised juveniles.
Power was high for all the blood parameters (>0.8, Table 2) for both
20% and 80% effect sizes.
The overall disappearance rate of juveniles from the beginning
to the end of the experiment was 39%. Neither treatment(b = "1.12, v21 = 2.30, P = 0.13) nor kangaroo sex (b = 0.21,
Contrary to our predictions, experimental removal of parasites
v21 = 0.08, P = 0.77) accounted for variation in disappearance.
in juvenile kangaroos had no effect on their body condition, mass
Mean (±SE) faecal egg counts of treated kangaroos (36 ± 24 epg)
gain or limb growth. Juvenile kangaroos treated with anthelmintics
were much lower than for control kangaroos (885 ± 251 epg,
had significantly higher albumin levels than control juveniles, but
F1,25.6 = 19.23, P < 0.001; Fig. 1) 12–33 days following treatment,
showed no differences in any other blood parameters. There was
resulting in a 99% reduction in faecal egg counts. There was no dif-
no evidence of parasite-induced mortality at our site, and the dis-
ference in the faecal egg counts within each group in either time
appearance rate of juveniles (39%) was comparable to that seen in
period (F1,3.1 = 2.41, P = 0.22). However, there was a significant
another eastern grey kangaroo population subject to predation by
interaction between time and treatment (F1,3.4 = 32.36, P = 0.007),
red foxes (Banks et al., 2000). Our results suggest that juvenile
such that faecal egg counts in the treated group increased more
eastern grey kangaroos are largely unaffected by gastrointestinal
than those in the control group. No cestode eggs were detected
parasitism in populations where burdens of G. trifidospicularis are
Author's personal copy
J. Cripps et al. / International Journal for Parasitology: Parasites and Wildlife 3 (2014) 88–94
Table 1Repeated measures ANOVA of treatment effects on selected ecological growth parameters for control and anthelmintic-treated juvenile eastern grey kangaroos at the AngleseaGolf Club, Victoria, Australia, from May to September 2012. The summaries show the sample size (n), the mean (±SE) increase (first to final capture) and the P-value for eachgroup. The statistical power of this experiment to detect a significant difference between treatment and control groups was calculated for effect sizes of 20% and 80%.
Body condition (scaled mass index)
Table 2Summary of the sample size (n), mean (±SE) (at the final capture) and test statistics of selected serum chemistry and haematological parameters for control and anthelmintic-treated juvenile eastern grey kangaroos at the Anglesea Golf Club, Victoria, Australia, from May to September 2012. The statistical power of this experiment to detect a significantdifference between treatment and control groups was calculated for effect sizes of 20% and 80%.
Total protein (g/L)
Red blood cells (!1012/L)
Haemoglobin (g/L)
Haematocrit (L/L)
* Denotes a significant difference between groups.
low, despite carrying high burdens of other gastrointestinal hel-
Arundel et al., 1990), perhaps explaining why we observed only
minth species.
an 8% difference in albumin levels between the two groups of
Defence against parasites incurs a cost, which may lead to
juveniles. The nematode M. baylisi may also feed on blood
resources being partitioned into immune response rather than
(Arundel et al., 1990), however clinical impacts have never been
growth (Zuk and Stoehr, 2002). As resources are limited, animals
confirmed. Larvae of a third species, R. rosemariae, can cause
undergoing rapid growth must partition and prioritize them
severe lesions on the gastric mucosa, yet hosts can carry large
appropriately (partitioning framework; Coop and Kyriazakis,
burdens without any obvious effects on health (Beveridge and
1999). Accordingly, a growing animal encountering parasites for
Presidente, 1978). As in all eastern grey kangaroo populations,
the first time would be expected to prioritise immunity over
helminth coinfection was ubiquitous in kangaroos at our site,
growth (Coop and Kyriazakis, 1999). We therefore predicted that
and juveniles were infected with between 5 and 8 different hel-
untreated (control) juvenile kangaroos, which were encountering
minth species (post-mortem data, Cripps, unpublished data).
nematode parasite larvae for the first time, would have a reduced
However, due to the inability to morphologically distinguish the
allocation to growth and energy reserves, and this would be
eggs of the various taxa in the faeces, it was impossible to deter-
reflected in their blood parameters. Field experiments such as ours
mine which combination of species were present in any individual
are rare in wildlife hosts, but have mostly shown significant sub-
in our study at any one time, and therefore hypoalbuminaemia
clinical effects on hosts (e.g. Hillegass et al., 2010; Stien et al.,
could not be attributed to any helminth species in particular.
2002). It was therefore surprising that the removal of parasites
Severe blood loss should also reduce concentrations of haemoglo-
had little effect on growth or the haematological variables exam-
bin and total protein, and lower the haematocrit, but we found no
ined, particularly as the faecal egg counts were high in the parasi-
differences in any of these despite our high statistical power. We
tised kangaroos. However, when compared to untreated controls,
are confident our experimental study would have detected even
anthelmintic-treated juveniles tended to gain more weight, and
small changes in any of the other blood parameters. Alternatively,
tended to have a longer leg and pes. Although not significant, the
the hypoalbuminaemia we observed could have resulted from
power analysis suggests that perhaps with larger sample sizes,
reduced food intake and consequent malnutrition, which is a com-
greater impacts of parasitism in juvenile grey kangaroos might
mon cause of reduced albumin synthesis (Rothschild et al., 1972).
have be seen. If individuals were investing extra energy and nutri-
Voluntary reductions in food intake are common during parasitic
ents in several areas at once (such as both skeletal and muscular
infections in livestock (Holmes, 1987; Kyriazakis et al., 1998), but
growth), it may have made it difficult to detect changes in each
whether parasite-infected kangaroos also exhibit anorexia is
parameter alone (Munger and Karasov, 1989), leading to low
unknown. Our study demonstrates that when levels of G. trifido-
spicularis are low, kangaroo hosts are able to tolerate their hel-
Haematological parameters can be altered by parasites directly
minth community and exhibit few subclinical effects from
through haematophagy (blood-feeding) and indirectly by limiting
the digestion and absorption of essential nutrients, such as amino
Increasing resource acquisition may be pivotal in allowing hosts
acids and protein (Colditz, 2008). Lowered albumin levels in kanga-
to reduce the potential costs of parasitism. Livestock hosts on high-
roos could be directly caused by blood loss and/or inflammation in
protein diets show reduced pathophysiological responses to para-
the gastrointestinal tract (Rothschild et al., 1988; Arundel et al.,
sitism (reviewed by Van Houtert and Sykes, 1996). For example,
1990). In juvenile eastern grey kangaroos, heavy infections of G.
infected sheep maintained on high protein diets increase their live
trifidospicularis have been associated with severe anaemia and clin-
weight gain by around 85% compared with control sheep (Van
ical disease (Arundel et al., 1990; I. Beveridge pers. obs), although
Houtert et al., 1995). Similarly, juvenile kangaroos could compen-
burdens (from post-mortems) at our site were more than five
sate for the costs of parasitism by using high-quality resources that
times lower than levels known to cause disease (400–1500,
offset nutrient and resource depletion. The best predictors of body
Author's personal copy
J. Cripps et al. / International Journal for Parasitology: Parasites and Wildlife 3 (2014) 88–94
condition in free-ranging kangaroos are the biomass and quality of
Arundel, J., Beveridge, I., Presidente, P., 1979. Parasites and pathological findings in
forage (Shepherd, 1987; Moss and Croft, 2009), and juvenile kanga-
enclosed and free-ranging populations of Macropus rufus (Demarest)(Marsupialia) at Menindee, New South Wales. Wildl. Res. 6, 361–379.
roos cannot sustain their growth on a poor-quality diet, even when
Arundel, J.H., Dempster, K.J., Harrigan, K.E., Black, R., 1990. Epidemiological
they are still suckling (Munn and Dawson, 2003, 2006). The climate
observations on the helminth parasites of Macropus giganteus (Shaw) in
at Anglesea is mild, and the fairways are irrigated, fertilized and
Victoria. Aust. Wildl. Res. 17, 39–51.
Banks, P., Newsome, A., Dickman, C., 2000. Predation by red foxes limits recruitment
regularly mown, which encourages new foliage with high protein
in populations of eastern grey kangaroos. Austral Ecol. 25, 283–291.
content (Jarman, 1974; Mattson, 1980). Furthermore, although
Beveridge, I., Presidente, P., 1978. Rugopharynx rosemariae new species (Nematoda:
the juveniles in our study should have been weaned, maternal care
Pharyngostrongylidae) from grey kangaroos (Macropus giganteus and M.
in kangaroos is a variable trait that can be influenced by a number
fuliginosus) with life cycle stages and associated pathology. Int. J. Parasitol. 8,379–387.
of factors, including a mother's age and/or body condition, and
Beveridge, I., Arundel, J.H., 1979. Helminth parasites of grey kangaroos, Macropus
environmental conditions (Stuart-Dick and Higginbottom, 1989).
giganteus (Shaw) and M. fuliginosus (Desmarest), in eastern Australia. Aust.
Differential maternal investment among individual mothers could
Wildl. Res. 6, 69–79.
Beveridge, I., Chilton, N.B., 2001. Co-evolutionary relationships between the
explain the high variability in the growth parameters we mea-
nematode subfamily Cloacininae and its macropodid marsupial hosts. Int. J.
sured, leading to a low power to detect change. The combination
Parasitol. 31, 976–996.
of resources from lactation and protein-rich pasture could have
Bordes, F., Morand, S., 2011. The impact of multiple infections on wild animal hosts:
a review. Infect. Ecol. Epidemiol. 1, 7346.
allowed infected juveniles to maintain their growth and body con-
Budischak, S.A., Jolles, A.E., Ezenwa, V.O., 2012. Direct and indirect costs of
dition in spite of parasitic infections.
co-infection in the wild: linking gastrointestinal parasite communities,
We have strong experimental evidence that juvenile kangaroos
host hematology, and immune function. Int. J. Parasitol. Parasites Wildl.
1, 2–12.
experience few subclinical effects from parasitism, contrary to our
Cameron, A., 2003. Reso for excel V.4.0 – Faecal egg count reduction test analysis.
predictions. While parasites clearly have subclinical effects in
AusVet Animal Health Services. University of Sydney.
many herbivorous hosts, data on free-ranging wildlife can be diffi-
Chiejina, S.N.C., Sewell, M., 1974. Worm burdens, acquired resistance and live
weight gains in lambs during prolonged daily infections with Trichostrongylus
cult to obtain and experimental field manipulations like ours are
colubriformis (Giles, 1892) Loos, 1905. Parasitology 69, 315–327.
imperative for investigating such relationships. Importantly, our
Colditz, I.G., 2008. Six costs of immunity to gastrointestinal nematode infections.
study is one of the first to combine both growth parameters and
Parasite Immunol. 30, 63–70.
haematological parameters, and supports the suggestion by
Coop, R., Kyriazakis, I., 1999. Nutrition–parasite interaction. Vet. Parasitol. 84, 187–
Budischak et al. (2012) that haematological parameters are more
Coulson, G., 2009. Behavioural ecology of red and grey kangaroos: Caughley's
sensitive to the subclinical effects of parasitism. In our study,
insights into individuals, associations and dispersion. Wildl. Res. 36, 57–69.
where levels of G. trifidospicularis were relatively low, gastrointes-
Cripps, J., Beveridge, I., Coulson, G., 2013. The efficacy of anthelmintic drugs against
nematodes infecting free-ranging eastern grey kangaroos, Macropus giganteus. J.
tinal helminths had minimal subclinical effects on the juvenile
Wildl. Dis. 49, 535–544.
eastern grey kangaroos. However, even small differences in growth
Erdfelder, E., Faul, F., Buchner, A., 1996. GPOWER: a general power analysis
and blood parameters could be biologically meaningful, and may
program. Behav. Res. Methods Instrum. Comput. 28, 1–11.
Green, A.J., 2001. Mass/length residuals: measures of body condition or generators
have implications for individuals later in life, particularly for life-
of spurious results? Ecology 82, 1473–1483.
history traits (Metcalfe and Monaghan, 2001). Future studies
Gulland, F.M., 1992. The role of nematode parasites in Soay sheep (Ovis aries)
should take a longitudinal, individual-based approach toward
mortality during a population crash. Parasitology 105, 493–503.
Gunn, A., Irvine, R.J., 2003. Subclinical parasitism and ruminant foraging strategies:
examining the cumulative effects of parasites over time.
a review. Wildl. Soc. Bull. 31, 117–126.
Hillegass, M.A., Waterman, J.M., Roth, J.D., 2010. Parasite removal increases
Conflicts of Interest
reproductive success in a social African ground squirrel. Behav. Ecol. 21, 696–700.
Holmes, P., 1987. Pathophysiology of nematode infections. Int. J. Parasitol. 17, 443–
There are no known conflicts of interest.
Hudson, P.J., Dobson, A.P., Newborn, D., 1992a. Do parasites make prey vulnerable
to predation? Red grouse and parasites. J. Anim. Ecol. 61, 681–692.
Hudson, P.J., Newborn, D., Dobson, A.P., 1992b. Regulation and stability of a free-
living host–parasite system: Trichostrongylus tenuis in red grouse. I. Monitoring
This work was supported by the Holsworth Wildlife Research
and parasite reduction experiments. J. Anim. Ecol. 61, 477–486.
Inwood, D., Catanchin, H., Coulson, G., 2008. Roo town slow down: a
Endowment. We thank Rachel Kane and staff at Anglesea Golf Club
community-based kangaroo management plan for Anglesea, Victoria. In:
for logistic support, and the many volunteers who assisted with
Lunney, D., Munn, A., Meikle, W. (Eds.), Too Close for Comfort:
fieldwork. Special thanks to Marco Festa-Bianchet and two anony-
Society of New South Wales, Mosman, Australia, pp. 1–8.
mous reviewers for perceptive comments on the manuscript,
Jarman, P.J., 1974. The social organisation of antelope in relation to their ecology.
Christine Andersen for technical assistance and to Rachel Sore
Behaviour 48, 215–267.
and Paul Carnell for statistical advice. This research was carried
Johnson, C.N., Jarman, P.J., Southwell, C.J., 1987. Macropod studies at Wallaby Creek
out with approval from The University of Melbourne's Animal Eth-
V. Patterns of defaecation by eastern grey kangaroos and red-necked wallabies.
Aust. Wildl. Res. 14, 133–138.
ics Committee (Project 1011709) and the Department of Sustain-
King, W.J., Wilson, M.E., Allen, T., Festa-Bianchet, M., Coulson, G., 2011. A capture
ability and Environment (research permit 10005557). There are
technique for free-ranging eastern grey kangaroos (Macropus giganteus)
no known conflicts of interest.
habituated to humans. Aust. Mammal 33, 47–51.
Kyriazakis, I., Tolkamp, B.J., Hutchings, M.R., 1998. Towards a functional explanation
for the occurrence of anorexia during parasitic infections. Anim. Behav. 56, 265–274.
Loyacano, A., Williams, J., Gurie, J., DeRosa, A., 2002. Effect of gastrointestinal
nematode and liver fluke infections on weight gain and reproductive
performance of beef heifers. Vet. Parasitol. 107, 227–234.
Mattson, W.J., 1980. Herbivory in relation to plant nitrogen content. Annu. Rev. Ecol.
Albon, S.D., Stien, A., Irvine, R.J., Langvatn, R., Ropstad, E., Halvorsen, O., 2002. The
Syst. 11, 119–161.
role of parasites in the dynamics of a reindeer population. Proc. R. Soc. Lond. B
McLeod, C., Wolff, J., 1968. Increased live weight gain and wool weight from
Biol. Sci. 269, 1625–1632.
anthelmintic drenching of ewe hoggets in South Canterbury. N.Z. J. Agric. Res.
Altizer, S., Nunn, C.L., Thrall, P.H., Gittleman, J.L., Antonovics, J., Cunningham, A.A.,
11, 407–419.
Dobson, A.P., Ezenwa, V., Jones, K.E., Pedersen, A.B., Poss, M., Pulliam, J.R.C.,
Mejia, M., Gonzalez-Iglesias, A., Diaz-Torga, G., Villafane, P., Formia, N., Libertun, C.,
2003. Social organization and parasite risk in mammals: integrating theory and
Becu-Villalobos, D., Lacau-Mengido, I., 1999. Effects of continuous ivermectin
empirical studies. Annu. Rev. Ecol. Evol. Syst. 34, 517–547.
treatment from birth to puberty on growth and reproduction in dairy heifers. J.
Anderson, N., Laby, R., Prichard, R., Hennessy, D., 1980. Controlled release of
Anim. Sci. 77, 1329.
anthelmintic drugs: a new concept for prevention of helminthosis in sheep. Res.
Metcalfe, N.B., Monaghan, P., 2001. Compensation for a bad start: grow now, pay
Vet. Sci. 29, 333–341.
later? Trends Ecol. Evol. 16, 254–260.
Author's personal copy
J. Cripps et al. / International Journal for Parasitology: Parasites and Wildlife 3 (2014) 88–94
Milner, J.M., Stien, A., Irvine, R.J., Albon, S.D., Langvatn, R., Ropstad, E., 2003. Body
Rothschild, M.A., Oratz, M., Schreiber, S.S., 1988. Serum albumin. Hepatology 8,
condition in Svalbard reindeer and the use of blood parameters as indicators of
condition and fitness. Can. J. Zool. 81, 1566–1578.
Scantlebury, M., Waterman, J., Hillegass, M., Speakman, J., Bennett, N., 2007.
Moore, I.T., Hopkins, W.A., 2009. Interactions and trade-offs among physiological
Energetic costs of parasitism in the Cape ground squirrel Xerus inauris. Proc. R.
determinants of performance and reproductive success. Integr. Comp. Biol. 49,
Soc. Biol. Sci. Ser., B 274, 2169.
Schmidt, R.L., Hibler, C.P., Spraker, T.R., Rutherford, W.H., 1979. An evaluation of
Moran, M.D., 2003. Arguments for rejecting the sequential Bonferroni in ecological
drug treatment for lungworm in bighorn sheep. J. Wildl. Manage. 43, 461–467.
studies. Oikos 100, 403–405.
Schulte-Hostedde, A.I., Zinner, B., Millar, J.S., Hickling, G.J., 2005. Restitution of
Moss, G., Croft, D., 2009. Body condition of the red kangaroo (Macropus rufus) in arid
mass-size residuals: validating body condition indices. Ecology 86, 155–163.
Australia: the effect of environmental condition, sex and reproduction. Aust. J.
Seitz, A., Ratte, H., 1991. Aquatic ecotoxicology: on the problems of extrapolation
Ecol. 24, 97–109.
from laboratory experiments with individuals and populations to community
Munger, J.C., Karasov, W.H., 1989. Sublethal parasites and host energy budgets:
effects in the field. Comp. Biochem. Physiol. C: Comp. Pharmacol. 100, 301–304.
tapeworm infection in white-footed mice. Ecology 40, 904–921.
Shepherd, N., 1987. Condition and recruitment of kangaroos. In: Caughly, G.,
Munn, A., Dawson, T., 2003. How important is milk for near-weaned red kangaroos
Shepard, N., Short, J. (Eds.), Kangaroos: Their Ecology and Management in the
(Macropus rufus) fed different forages? J. Comp. Physiol. B Biochem. Syst.
Sheep Rangelands of Australia. Cambridge University Press, Cambridge, pp.
Environ. Physiol. 173, 141–148.
Munn, A.J., Dawson, T.J., 2004. The ecophysiology of survival in juvenile red
Stien, A., Irvine, R.J., Ropstad, E., Halvorsen, O., Langvatn, R., Albon, S.D., 2002. The
kangaroos Macropus rufus: greater demands and higher costs. Aust. Mammal 26,
impact of gastrointestinal nematodes on wild reindeer: experimental and cross-
sectional studies. J. Anim. Ecol. 71, 937–945.
Munn, A.J., Dawson, T.J., 2006. Forage fibre digestion, rates of feed passage and gut
Stuart-Dick, R.I., Higginbottom, K.B., 1989. Strategies of parental investment in
fill in juvenile and adult red kangaroos Macropus rufus Desmarest: why body
Macropodoids. In: Grigg, G., Jarman, P., Hume, I. (Eds.), Kangaroos. Wallabies
size matters. J. Exp. Biol. 209, 1535–1547.
and Rat-Kangaroos. Surrey Beatty & Sons Pty Limited, Chipping Norton,
O'Kelly, J.C., Post, T.B., Bryan, R.P., 1988. The influence of parasitic infestations on
Australia, pp. 571–592.
metabolism, puberty and first mating performance of heifers grazing in a
Sykes, A.R., 1987. Endoparasites and herbivore nutrition. In: Hacker, J., Ternouth, J.
tropical area. Anim. Reprod. Sci. 16, 177–189.
(Eds.), Nutrition of Herbivores. Academic Press, New South Wales, pp. 211–232.
Peig, J., Green, A.J., 2009. New perspectives for estimating body condition from
Van Houtert, M., Barger, I., Steel, J., 1995. Dietary protein for young grazing sheep:
mass/length data: the scaled mass index as an alternative method. Oikos 118,
interactions with gastrointestinal parasitism. Vet. Parasitol. 60, 283–295.
Van Houtert, M.F.J., Sykes, A.R., 1996. Implications of nutrition for the ability of
Poole, W., Carpenter, S., Wood, J., 1982. Growth of grey kangaroos and the reliability
ruminants to withstand gastrointestinal nematode infections. Int. J. Parasitol.
of age determination from body measurements. I. The eastern grey kangaroo
26, 1151–1167.
Macropus giganteus. Aust. Wildl. Res. 9, 9–20.
Watson, M.J., 2013. The costs of parasites – What drives population-level effects?
Poole, W.E., 1975. Reproduction in the two species of grey kangaroos, Macropus
Meta-analysis meets life-history. Int. J. Parasitol. Parasites Wildl. 2, 190–196.
giganteus Shaw and M. fuliginosus (Desmarest). II. Gestation, parturition and
Wood, I., Amaral, N., Bairden, K., Duncan, J., Kassai, T., Malone, J., Pankavich, J.,
pouch life. Aust. J. Zool. 23, 333–353.
Reinecke, R., Slocombe, O., Taylor, S., 1995. World Association for the
Poole, W.E., 1983. Breeding in the grey kangaroo, Macropus giganteus, from
Advancement of Veterinary Parasitology (WAAVP) of guidelines for evaluating
widespread locations in eastern Australia. Wildl. Res. 10, 453–466.
the efficacy of anthelmintics in ruminants (bovine, ovine, caprine). Vet.
Riney, T., 1955. Evaluating condition of free-ranging red deer (Cervus elaphus), with
Parasitol. 58, 181–213.
special reference to New Zealand. NZ J Sci Technol 36, 429–463.
Zuk, M., Stoehr, A.M., 2002. Immune defense and host life history. Am. Nat. 160, 9–
Rothschild, M.A., Oratz, M., Schreiber, S.S., 1972. Albumin synthesis. New Eng. J.
Med. 286, 748–757.
Source: http://www.angair.org.au/images/stories/angair/kangaroos/parasites.pdf
TABLE OF CONTENTS *Perspectives from Biogen leadership For the fourth consecutive year, our global colleagues participated in ON THE COVER Biogen's Care Deeply Volunteer Day, which shattered last year's record Inspired by the new Biogen turnout with a 54-percent increase in employee participation. This worldwide logo, the front cover graphic
Ganadería Colombiana Sostenible Servicios ambientales que proveen los Sistemas Silvopastoriles y los beneficios para la biodiversidad Andrés Felipe Zuluaga S. Carolina Giraldo E. Julián David Chará. Fotografía Portada: David Fajardo. Andrés Felipe Zuluaga S. Carolina Giraldo E. Adolfo Galindo.