Doi:10.1016/j.pbb.2006.12.00

Pharmacology, Biochemistry and Behavior 86 (2007) 189 – 199
Adolescent cortical development: A critical period
of vulnerability for addiction
Fulton Crews ⁎, Jun He, Clyde Hodge
Bowles Center for Alcohol Studies, School of Medecine, University of North Carolina at Chapel Hill, NC 27599, United States
Received 24 July 2006; received in revised form 20 November 2006; accepted 4 December 2006
Available online 12 January 2007
Cortical growth and remodeling continues from birth through youth and adolescence to stable adult levels changing slowly into senescence.
There are critical periods of cortical development when specific experiences drive major synaptic rearrangements and learning that only occurduring the critical period. For example, visual cortex is characterized by a critical period of plasticity involved in establishing visual acuity.
Adolescence is defined by characteristic behaviors that include high levels of risk taking, exploration, novelty and sensation seeking, socialinteraction and play behaviors. In addition, adolescence is the final period of development of the adult during which talents, reasoning andcomplex adult behaviors mature. This maturation of behaviors corresponds with periods of marked changes in neurogenesis, cortical synapticremodeling, neurotransmitter receptors and transporters, as well as major changes in hormones. Frontal cortical development is later inadolescence and likely contributes to refinement of reasoning, goal and priority setting, impulse control and evaluating long and short termrewards. Adolescent humans have high levels of binge drinking and experimentation with other drugs. This review presents findings supportingadolescence as a critical period of cortical development important for establishing life long adult characteristics that are disrupted by alcohol anddrug use.
2006 Elsevier Inc. All rights reserved.
Keywords: Alcohol; Adolescence; Cortical development; Binge drinking; Critical period
Introduction: Adolescence; a unique period of development . . . . . . . . . . . . . . . . . . .
Neurotransmitter systems and adolescent development . . . . . . . . . . . . . . . . . . . .
Transcription factor CREB and growth factor BDNF in adolescence . . . . . . . . . . . . . . . .
Adolescent behavior: Risky, motivated, and vulnerable . . . . . . . . . . . . . . . . . . . .
⁎ Corresponding author. Bowles Center for Alcohol Studies, The University of North Carolina at Chapel Hill, 1021 Thurston Bowles Building, CB 7178, Chapel
Hill, NC 27599-7178, United States. Tel.: +1 919 966 5678; fax: +1 919 966 5679.
E-mail address: (F. Crews).
URL: (F. Crews).
0091-3057/$ - see front matter 2006 Elsevier Inc. All rights reserved.
F. Crews et al. / Pharmacology, Biochemistry and Behavior 86 (2007) 189–199
Binge drinking during critical periods in cortical development may lead to life long changes of executive function . . . . . 195
1. Introduction: Adolescence; a unique period of
of such synaptic changes are not well known, it is speculated
that such remodeling is the biological basis of developmentalplasticity where the neurological circuits are effectively shaped
Adolescence is a critical period of development during the
to adapt to the environmental needs leading to mature adult
transition from childhood to adulthood. The ages associated
behavior. Such a period of remodeling could also make the
with adolescence are commonly considered in humans to be
adolescents more vulnerable to external insults and other
approximately 12 to 20–25 years of age, and postnatal days
(PND) 28 to 42 ) in rodents. Adolescence is best
The prefrontal cortex (PFC) and the limbic system, which
defined by characteristic adolescent behaviors that include high
includes the hippocampus, amygdala, nucleus accumbens
levels of risk-taking, high exploration, novelty and sensation
(NAc), prefrontal, frontal and orbital frontal cortices and the
seeking, social interaction, high activity and play behaviors that
hypothalamus, undergo prominent reorganization during ado-
likely promote the acquisition of the necessary skills for
lescence. Absolute PFC volume declines in adolescence in
maturation and independence (). Adolescent
humans (as well as in rats
behaviors are shared across species, for example, high social
Substantial loss of synapses, especially the
interactions are found in human adolescents (increased
excitatory glutamatergic inputs to the PFC, occurs during the
communication with peers and increased number of conflicts
adolescent period in humans and nonhuman primates (
with parents) () as
In contrast to such
well as in adolescent rodents (increased peak level of play
adolescent-associated pruning, dopamine and serotinin (5-HT)
behavior and affiliative behaviors like huddling, grooming etc.)
inputs to PFC increase during adolescence to peak levels well
above those seen earlier or later in life (
These behaviors have been suggested to help adolescents
). Cholinergic innervation of PFC
develop the social skills needed when they become independent
also increases in adolescence to reach mature levels in rats
from their family or become senior adults in their group. In
) and humans (). In the
rodents, increased social interaction helps guide their food
hippocampus, the exuberant outgrowth of excitatory axon
choices () and other adult behaviors such as sexual
collaterals and synapses during youth are morphologically
and aggressive behaviors (Unfor-
remodeled and branches within dendritic arbors are pruned
tunately, these high levels of novelty/sensation-seeking beha-
during adolescent maturation ). Similarly,
viors are also strong predictors of drug and alcohol use among
significant dendritic pruning and synaptic regression occur in
medial amygdala (), nucleus accumbens (NAc)
). This review will cover brain maturation of neuroanat-
and the hypothalamus
omy, neurotransmission and behavior during adolescence and
during adolescence.
present the postulate that the adolescent brain is a critical period
Although most synaptic pruning is likely glutamatergic, dopa-
of vulnerability for disruption of brain regions important for
mine receptor expression peaks in early adolescence (PND28)
followed by a one-third loss of receptors during PND35 toPND60 In terms of hypothalamic func-
2. Adolescent brain remodeling
tion, adolescent rats often exhibit more prolonged stress-in-duced increases in cortisol than adults (In
The adolescent brain is in a unique state of transition as it
addition, rats at PND 28 were found to show less stress-induced
undergoes both progressive and regressive changes providing
Fos-like immunoreactivity in cortical and amygdaloid nuclei
a biological basis for the unique adolescent behaviors and the
than adult rats (but higher novelty-in-
associated changes in behavior during maturation to adult-
duced Fos activation in hippocampus during this period
hood. Human magnetic resonance imaging (MRI) studies have
). Thus, environmental alterations in gene transcrip-
demonstrated an inverted U-shape change in the gray matter
tion are unique during adolescence and likely impact the active
volume during adolescent period, with a pre-adolescent
remodeling of synaptic connections. The following paragraphs
increase followed by a post-adolescent decrease
review neurochemical markers of adolescent brain remodeling
At the cellular levels, these changes
to illustrate the high plasticity of normal brain development.
correspond with the marked overproduction of axons and
The remodeling of the adolescent brain may represent a critical
synapses in early puberty, and rapid pruning in later
period of development during which alcohol and drugs may
become significant environmental factors modulating brain
). Although the exact mechanisms
F. Crews et al. / Pharmacology, Biochemistry and Behavior 86 (2007) 189–199
3. Critical periods of cortical development
cortical synaptic pruning and myelination Inhibitory control involves executive func-
Critical periods are specific windows during development
tions that improve from adolescence to adulthood. Studies
when both genetic driven processes and environmental process-
measuring behavioral inhibition on a Go-No-Go task and fMRI
es, e.g. nature and nurture, interact to establish functional
data reveal greater activation of dorsolateral frontal cortex and
characteristics. These interactions correspond to structural
orbitofrontal cortexes in children, than adolescence, and greater
rearrangements of the cerebral cortex that occur during this
in adolescence than adults with the adults showing the lowest
specific developmental window. As described above, cortical
dorsolateral, but equal orbitofrontal activation and greater
development in humans occurs over the first 3 decades of life
inhibitory control performance (
with grey matter changes correlating with post-mortem findings
). These studies support the concept that the immature
of brain regionally different synaptic pruning and myelination
brain with excess synapses causes more extensive and less
during the transitions from childhood to adolescence to
efficient frontal activation and lower performance compared to
adulthood. The human visual cortex reaches a peak of synaptic
adults that have a more pruned and myelinated frontal cortex that
overproduction around the 4th month after birth followed by
results in more focused, lower overall activation and faster
synaptic elimination starting after that and continuing until
reaction times and better performance
preschool age at which time synaptic density stabilizes to adult
Taken together these studies suggest that remodeling
levels Other cortical areas develop at
of the cortex during the transitions from youth to adolescence to
different ages with dorsal parietal and primary sensorimotor
adulthood has functional implications for the entire adult stages
regions showing grey matter loss at ages 4–8, and parietal areas
of language and spacial orientation changing around ages 11–13
Environmental plasticity of visual cortex development has
and prefrontal areas involved in integrating information from
been extensively studied. Hubel and Wiesel found that depriving
senses, reasoning and other "executive functions" maturing last
an eye of light altered cortical responses to light only if the
during late adolescence
deprivation occurred during a "critical period" of cortical
These age-related changes in cortical structure involve improved
development. The critical period for visual cortical plasticity is
function. Cortical thinning in the left dorsal frontal and parietal
defined as the period during which monocular deprivation
lobes correlates with improved performance on a test of general
(covering one eye) results in a shift in cortical neuronal spiking
verbal intellectual functioning between the ages of 5 to 11
responses away from the covered eye and increased spiking
(Other studies following individuals from
responses to the active eye. The spiking responses are shifted to
age 6 through 19 found that individuals with superior
the open eye only if the deprivation occurs during the critical
intelligence show the greatest changes in frontal cortical
period. The critical period for visual cortex plasticity in rodents
thickness compared to individuals with high or average
overlaps with early adolescence being between PD19 and PD 32
intelligence ). These changes likely are a
). Studies indicate that an activity
combination of environment and genetic regulation of cortical
dependent synaptic plasticity occurs during this critical period
development and overall function. Environmental experiences
that allows cortical adjustments to environmental factors. Dark
and training are known to induce changes in cerebral cortex
rearing delays the critical period of visual cortex development
including neurochemical, altered cortical thickness, size of
into adulthood likely by decreasing BDNF expression and
synaptic contacts and dendritic structure as well as improving
GABA synaptic strength Genetic BDNF
performance on learning tests
overexpression or benzodiazepines administration can acceler-
Learning in humans during studying for exams (
ate the appearance of the critical period of visual cortical
or practicing juggling ) alters
development. Environment and genetic programming interact to
cortical structure consistent with environment contributing to
regulate synaptic organization during the critical period
structural changes in brain. These developmental processes are
resulting in a mature cortex. The mature cortex is stable and
thought to underlie time-limited windows when specific
does not undergo the dramatic shift in cortical synaptic plasticity
experiences can drive development, e.g. critical periods of
with monocular deprivation or dark rearing. Thus, early
plasticity or learning that can only occur during these critical
adolescence is a critical period for visual cortical maturation.
period windows (For example, learning a
Ethanol treatment has been shown to lead to a permanent
second language is thought to be optimal during a critical period
impairment of visual neocortex plasticity and to be particularly
of development (The complexity
toxic to adolescent brain. Studies in ferrets have found that
of higher brain functions makes it difficult to relate synaptic
ethanol treatment before the critical period prevents ocular
rearrangements to alterations in function. The synaptic rearran-
dominance from occurring in neurons during the critical period
gements and increased myelination of frontal cortical areas in
mid to late adolescence could be involved in altered executive
iological single unit recording indicates that alcohol exposure
functions. Behavioral studies show that performance on tasks
weakened neuronal orientation selectively while preserving
including inhibitory control, decision making and processing
robust visual responses Further, optical
speed continues to develop during adolescence. During
imaging maps of intrinsic synaptic signaling from the eyes to the
adolescence tasks of selective attention, working memory and
visual cortex are highly contrasted in controls following the
problem solving improve consistently correlating with frontal
critical period of synaptic ocular dominance, but have little
F. Crews et al. / Pharmacology, Biochemistry and Behavior 86 (2007) 189–199
contrast or no ocular dominant columns after ethanol adminis-
a major contribution of cortical remodeling. In adolescent
tration. Disruption of ocular dominant columns in visual cortex
monkeys significant changes occur in pre- and post-synaptic
is known to reduce visual acuity and monocular deprivation
markers of GABAergic synapses in the prefrontal cortex during
during the critical period essentially creates a blind eye due to
adolescence ). Generally, basal levels of
altered synaptic connections These
GABAA receptor-mediated chloride uptake are greater in the
studies indicate that alcohol exposure preceding and during
cortex of adolescents than that of adults ()
critical periods of cortical development disrupt cortical devel-
and the responsiveness of cortical GABAA neurotransmission to
opment resulting in reduced function permanently. Studies of
stressors decreases from adolescence to adulthood (
binge drinking induced brain damage in rats have found that
In hippocampus, the expression of GABA transporter 1
adolescent forebrain is particularly sensitive to ethanol induced
(GAT-1) and glutamic acid decarboxylase (GAD), e.g. GABA
neurodegeneration ). Further, adolescent
synthase, peak around early infancy
brain has been found to be particularly sensitive to ethanol
although hippocampal GABAB receptor regulation of
induced inhibition of neurogenesis, the formation of new
synaptic transmission does not mature until adolescence
neurons ). These studies indicate that ethanol
(PND35) (). GABAA receptors are
disrupts cortical remodeling during critical periods and is
heterogeneous group of receptors composed of two alpha, one
particularly neurotoxic to the adolescent brain.
beta and two delta or other subunits. There are multiple alpha
Ethanol is known to interact with glutamatergic NMDA
subunits that can contribute to GABAA receptors and varied
receptors, GABAa receptors, DA pathways, CREB transcrip-
subunits modulate GABAA receptor function and responsive-
tion, and neurogenesis. Each of these systems is undergoing
ness to GABA, anti-anxiety drugs, neurosteroids, alcohol and
extensive remodeling in adolescence. The following sections
other modulators of GABA transmission. In fact, the subunit
review the remodeling of these neurochemical markers known
composition of GABAA receptor subunits largely determines the
to be sensitive to ethanol.
pharmacological and electrophysiological properties of GABAneurotransmission
4. Neurotransmitter systems and adolescent development
GABAA subunits undergo dramatic postnatal reorgani-zation ). The alpha1 subunit that contributes to
4.1. Glutamate and NMDA receptor systems
sedative, amnestic and anticonvulsant actions of GABA under-goes a dramatic increase in the frontal cortex between youth (10
The binding of cortical glutamate to its NMDA receptor
PND) and adolescence (30 PND) followed by a significant de-
subtype peaks in early adolescence, and declines significantly
cline in the transition to adulthood (90 PND) becoming rela-
thereafter, with a loss of one-third of NMDA receptors by
tively stable at later ages This likely represents
PND60 (). Such synaptic
remodeling of GABAergic synapses. Alpha2 and Alpha3
pruning contributes to the loss of excitatory glutamate input to
subunits show a similar increase from youth to adolescence, in
NAc (and a reduction in
frontal cortex, however, alpha2 stays elevated through adult-
accumbal NMDA receptors
hood and alpha3 GABAA subunits decline slowly reaching
during adolescent brain maturation. Interestingly, long-term
statistical significance at 9 months. Alpha5 GABAA subunits are
potentiation (LTP), a measurable increase in synaptic strength
high in youth and in different brain regions decline through
and a form of neuroplasticity, is more frequently found in NAc of
adolescence into adulthood. Alpha2, 3 and 5 subunits contribute
adolescent mice compared to adults consistent with adolescence
to anxiolytic and other properties of GABA modulators
being a highly plastic period of mesolimbic brain development
(Fritschy and Brunig, 2003). Interestingly, the alpha2 subunit
(This plasticity was also demonstrated in
of the GABAA receptor complex, whose expression steadily
other limbic regions that are believed to be involved in drug
increases across the brain from early youth throughout
addiction, including amygdala (VTA
adolescence has been implicated in the genetics
(and hippocampus ).
of alcoholism. A sequence difference in the alpha2 subunit of the
Alcohol and other drugs are known to modulate glutamatergic
GABAA receptor has been found to be about twice as common in
transmission and alter limbic brain development (
alcohol dependent German patients than matched controls
). Therefore, glutamate and NMDA receptor systems play a
consistent with contributing to genetic predisposition to
crucial role in the neurochemical remodeling in adolescents,
alcoholism (Interestingly, chronic alcohol in
especially in limbic brain regions that are highly plastic and
adults decreases cerebral cortical alpha2 and alpha3 subunits
actively undergoing remodeling. The impact of alcohol and
resulting in loss of benzodiazepine recognition sites and likely
other illicit drugs on the development of glutamatergic synapses
changing overall GABA transmission
in these brain regions is critical for understanding the particular
Studies in adolescents have not been done, however, alcohol
vulnerability of adolescents to drug addiction.
induced changes in GABA subunits during the active period ofcortical development might result in differential stabilization of
4.2. GABAergic systems
adult GABAA receptor synaptic organization that remaindisrupted for long periods in adulthood. The maturation of
GABAergic (γ-aminobutyric acid) neurotransmission, as the
GABAergic neurotransmission from infancy to adolescence to
major inhibitory neurotransmitter in the brain, that likely makes
adulthood likely contributes to inhibitory interneuron fine-
F. Crews et al. / Pharmacology, Biochemistry and Behavior 86 (2007) 189–199
tuning of synaptic inputs improving discrimination of signals
expressed in humans, cats and rodents at birth, but decline
and more efficient processing. Disruption by alcohol and other
dramatically during adolescence
addictive drugs during the adolescent remodeling of GABA-
mediated inhibitory control of neuronal circuitry could alter
susceptibility to alcohol dependence and other drug addiction in
NAc is reported to be approximately 4-fold lower in adolescent
rats (PND30-40) than either younger rats (PND10-15) or matureadults (PND60-80) ). Interestingly, low 5-HT
4.3. Dopaminergic systems
activity in adolescence has been suggested contribute tocommon adolescent behaviors such as hypersensitivity to
Dopaminergic transmission contributes to attention, reward,
mild stressors, increased anxiety and alcohol drinking
movement, hormone regulation and multiple other important
In contrast to the alteration in serotonin
physiological processes. Postnatal reorganization of dopami-
receptors during adolescence, serotonin transporters steadily
nergic neurotransmission is brain regional and receptor subtype-
increase from PND7 to adulthood without significant pruning in
specific. In rat frontal cortex, entorhinal cortex and hippocam-
striatum and NAc Studies modeling
pus, dopamine D1, D2 and D4 receptors rise several fold from
adolescence binge drinking in rats have found marked increases
PND7 to PND35, e.g. adolescence, and then stabilize to
in adult levels of serotonin transporters ().
adulthood ). In striatum and
Thus, serotonin neurotransmission undergoes dramatic remo-
nucleus accumbens (NAc) dopamine receptors are overpro-
deling from youth through adolescence into adulthood and it is
duced with subsequent pruning of approximately one-third
sensitive to alcohol and drug disruption.
during adolescent suggesting maturational remodeling of motor
In summary, major neurotransmitter systems are not mature
and reward pathways (
at birth and postnatal brain development continues through
Furthermore, dopamine D3 receptors do
adolescence, with remodeling most pronounced in frontal and
not reach peak levels until adulthood (PND60) in striatum, NAc
limbic regions.
and olfactory tubercle ). In contrast,dopamine transporters increase 6–7 fold steadily throughout
5. Transcription factor CREB and growth factor BDNF in
brain from PD7 to PD60 in striatum in contrast to the
remodeling of dopamine receptors in adolescent striatum(). This substantial postnatal remodeling of
The cAMP-response element binding protein (CREB) is an
dopamine neurotransmission during adolescence may contrib-
important mediator for the differentiation and maturation of
ute to a stabilization of behaviors established during adoles-
CNS neurons. CREB is also critical for induction of trophic
cence. Alcohol and drug taking may alter the maturation of
factors such as BDNF, for neuronal vitality and for learning and
dopamine neurotransmission during adolescence contributing
to altered development of attitudes, actions and social rewards.
phosphorylation, CREB is activated to propagate signals fromsynapses to the nucleus to the expression of genes necessary for
4.4. Serotonergic systems
synaptic plasticity (). As such a key transcriptional factor for
Serotonergic neurotransmission undergoes reorganization
neuronal growth, it is not surprising that the expression of CREB
during postnatal development and is important for mood, sleep,
can play a critical role in postnatal neurochemical remodeling.
anxiety and many other complex behaviors. In humans and rats,
Phospho-CREB (pCREB), the transcriptionally active form of
5-HT neurons are generated prenatally
CREB, is highly expressed in early postnatal development
) with brain 5-HT levels peaking early in
(PND7) and declines during adolescence to adult levels in both
life, then decreasing to adult levels (
hippocampus and cortex (Additionally,
Postnatal reorganization of developing
CREB activation occurs prior to the expression of BDNF (brain-
serotonergic projections is exemplified by fluctuations of the
derived neurotrophic factor) and neurotrophin-3 (NT3) (
number of serotonergic synapses during this period, 5HT
, consistent with CREB being upstream and
synapses reach adult levels at PND14 and then in rat basal
activating transcription of these neurotrophic factors. BDNF is
forebrain drop to significantly lower levels during early
involved in the regulation of neuronal differentiation, survival
adolescence (). The
and neuroplasticity as well as being linked with a variety of
reorganization of 5-HT receptor expression is also pronounced
neurological and psychiatric disorders (epilepsy, mood disorder,
during development, likely relating to reorganization of
bipolar depression) in children, adolescents as well as adults
serotonergic innervation patterns. For example, 5-HT2A recep-
tors reach cortical peak expression just before adolescence and
CREB and BDNF interact in a variety of brain regions and are
then progressively decline to adult levels correlated with
known to play a critical role in addition ).
increased innervation and pruning of 5-HT axons in rat and
Thus, the developmental alterations in CREB-BDNF contribute
monkey (Similarly, 5-HT7
to continuous modeling of brain through youth, adolescence into
receptors exhibit transient expression patterns in striatum and
adulthood and are vulnerable to alcohol and drug induced
hippocampus (5-HT1A receptors are highly
disruption of development.
F. Crews et al. / Pharmacology, Biochemistry and Behavior 86 (2007) 189–199
6. Neurogenic processes in adolescent brains
into action (). Increased adolescentreward sensitivity, motivation and action is likely related to the
Although neurogenesis is primarily an early developmental
synaptic remodeling of striatal, limbic and frontal brain regions
process with most neurons being formed in the prenatal and
as the final stage of development to adulthood.
early postnatal periods, it continues into adulthood within
In addition to reward circuits, a variety of other neural
specific adult brain regions including the forebrain subven-
systems also exhibit developmental changes during adolescence
tricular zone (SVZ) and hippocampal dentate gyrus (DG) where
that lead to behavioral and cognitive alterations. For example,
neurogenesis continues into senescence. Generating and
evidence suggests that a variety of self-regulatory executive
integrating new neurons into preexisting neuronal circuits is
functions are still maturing during adolescence. For this reason,
believed to enable the hippocampus to adapt to novel and more
adolescents are in the sometimes unfortunate situation of having
complex situations (The contribution of
poor judgment and lack of impulse control even though they are
adult hippocampal neurogenesis to learning and memory (
driven to seek increasing levels of novelty and external stimu-
as well as mood and affective state (
lation. A variety of brain systems mediates impulse control
) is supported by many studies. Adolescent neurogenesis
) but maturation of prefrontal cortical
and its role in the brain remodeling and unique adolescent
systems appears to track development of executive functions
behaviors have not been investigated. Studies have indicated
that adolescents have higher levels of neurogenesis in the
Adolescence is also characterized by the appearance of
hippocampus and adolescent brain
strong emotional states where some individuals experience
neurogenesis is very sensitive to alcohol-induced inhibition
striking changes in mood that are sometimes difficult to
(). Disruption of neurogenesis by drugs and
distinguish from clinical syndromes, such as depression
alcohol during adolescence could produce long-lasting changes
An understanding of mood
in adulthood.
changes during adolescence may be gained from evaluatingneurobiological systems that regulate adult mood disorders.
7. Adolescent behavior: Risky, motivated, and vulnerable
Along these lines, one potentially important outcome ofdevelopmental changes in gene transcription (e.g., CREB) and
As just noted, the adolescent brain undergoes remodeling in
growth factor (e.g., BDNF) expression is altered neurogenesis
a variety of structural and functional regions, particularly
(see above). It has been hypothesized that decreased neurogen-
corticolimbic and frontal regions known to regulate emotional
esis in the hippocampus is a mechanism underlying mood
as well as analytical and executive processes. Simultaneous
changes, such as depression (
with these changes, adolescents demonstrate new behaviors that
which are common during adolescence. Although the
are associated with acquisition of adult cognitive and emotional
functional consequences of altered neurogenesis to adolescent
repertoires (These are normal adaptive changes
behavioral changes remain to be studied, some noteworthy
that help usher the adolescent into adulthood. However,
linkages exist. For example, stress and elevated stress
adolescents also exhibit increased health-risk behaviors that
hormones, which are hallmarks of adolescence, alter neurogen-
represent the leading causes of morbidity and mortality among
esis and precipitate changes in mood
the adolescent age group (). Disruption of
adolescent development by environmental factors, particularly
Antidepressant drugs that target serotonin and
alcohol and other abused drugs might produce subtle changes
norepinephrine systems, both of which change during adoles-
other than the known pronounced mortality that have a delayed
cence, increase hippocampal neurogenesis (
impact on the quality of adult life. For this reason, it is important
When taken together
to understand adolescent behavior and neurobiology in the
with evidence that antidepressants also increase serotonin,
context of the developing brain in order to appreciate the
CREB and BDNF activity ), these
adaptive changes within a critical period of acquisition of adult
data suggest that developmental changes in these pathways may
cognitive and emotional repertoires.
underlie some of the mood alterations that characterize
One of the most pronounced changes in adolescent behavior
adolescence. In addition, evidence shows that enriched
is the characteristic increased risk taking. Epidemiological
environment and exercise are associated with increased mood
studies show that human adolescents engage in more risky
and neurogenesis, whereas stress is associated with depression
behavior, which includes hazardous driving, unprotected sex,
and decreased neurogenesis (
and substance abuse as compared to adults (
). The adolescent trait of impulsivity is also
These behaviors are associated with low levels of
normalized by enriched rearing during adolescence (
anxiety regarding the potential for harm
). Moreover, adolescents who have enriched envi-
). One explanation for age-related differences in risk taking
ronments in the form of socially active friends and engaged
is the characteristic reduction in reward sensitivity that leads
parents exhibit fewer behavioral problems and engage in less
adolescents to seek higher levels of novelty and external
risky behavior ). Thus, it is plausible that
stimulation. Changes in reward sensitivity may reflect matura-
both developmentally and environmentally regulated changes in
tional differences in mesolimbic neural circuits (
neurogenesis may underlie alterations in mood and behavior
which regulate the translation of motivation
that accompany adolescence.
F. Crews et al. / Pharmacology, Biochemistry and Behavior 86 (2007) 189–199
Overall, adolescents exhibit a variety of behavioral changes
9. Binge drinking during critical periods in cortical
that reflect normal development of brain systems. Paradoxical-
development may lead to life long changes of executive
ly, these developmental changes also create conflicts in
behavioral repertoires that mark the unique vulnerability ofthis developmental period. A few of these have been discussed
The effects of alcohol on adolescent brains are different from
above in terms of specific systems that may regulate specific
those on adults. Adolescents are less sensitive to the sedative
behaviors. Ultimately, however, a more complete understanding
effects of alcohol (which allows them
of the paradoxical properties of adolescent behavior is likely to
to binge drink, however, they are more vulnerable to alcohol-
require a multidisciplinary systems approach that integrates
induced neurotoxicity ). The increase in
developmental, behavioral, and neural analyses.
sensitivity of the adolescent brain to toxicity and the dynamicsynaptic remodeling of the maturing adolescent brain may
8. Adolescent alcohol abuse is common
enhance the strong learning components of heavy drinkingbehaviors and the loss of important self-control and goal setting
Alcohol use among adolescents is common. As described
components of the maturing brains executive centers. Indivi-
earlier, adolescent high risk-taking, thrill and novelty-seeking
duals who start drinking before the age of 15 are four times
behaviors promote heavy drinking and other drug experimen-
more likely to become alcohol dependent at some time in their
tation. Individuals in their teens and early twenties are among
life (Current studies indicate that
the heaviest episodic drinkers. For example, among U.S. high
25–35% of high school students began drinking before the age
school students 12% of 8th graders (13–14 years of age), 22%
of 13 ). Studies of adolescent individuals with
of 10th graders and 28% of 12th grade seniors reported heavy
alcohol use disorder have found smaller prefrontal grey and
episodic drinking within the past 2 weeks
white matter volumes than age matched controls. Lower
According to the National Institute of Drug Abuse, 82%
prefrontal volumes correlated with a higher maximum number
of adolescents have tried alcohol by the time they reach their
of drinks per drinking episode (It is
senior year in high school. For college students 44% binge drink
likely that both genetics and environment (heavy drinking)
every two weeks and 19% are frequent binge drinkers, having
contribute to the alcohol use disorder and lower prefrontal
more than 3 binge drinking episodes per week (
volumes in adolescence with alcohol use disorder. Studies of
Thus, adolescents are often drinking large quantities of
social drinkers have found that the heaviest binge drinkers have
more negative moods and performed worse on executive
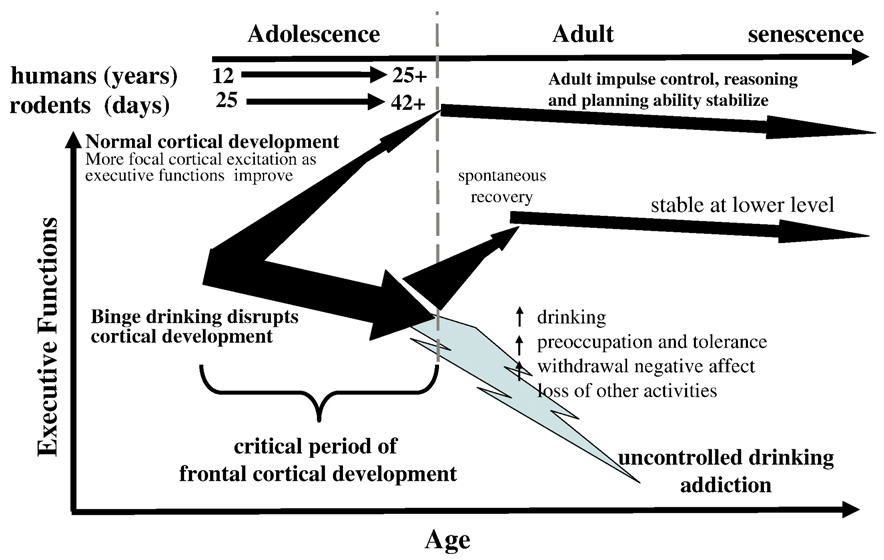
Fig. 1. Adolescent alcohol abuse disrupts frontal cortical development and maturation of executive function. This schematic diagram emphasizes the normal focusingof cortical excitation during cerebral tasks that occurs during the transition from adolescent to adult as indicated by the upward narrowing arrow. Frontal corticalremodeling is associated with improved performance at tasks and stabilization of reasoning, impulse control, goal setting, maturation of risk taking, reward sensitivity,motivation, and emotional states. Adult executive functions stabilize after adolescence with a slow decline in senescence. Individuals who have talent (genetics) andtraining (environment) are most likely to achieve their best abilities at the juncture of adulthood. Individuals who binge drink during adolescence damage and disruptforebrain cortical development during a critical period of behavioral and cortical maturation. Binge drinking interferes with cortical remodeling as illustrated by thelarge vertical slightly downward arrow that does not focus. Following adolescence many individuals spontaneously or through therapy reduce their drinking andpartially recover executive functions, although they remain lower in ability than those who develop normally. Other individuals remain heavy drinkers continuing todrink. Through the life course many will escalate drinking due to stress, tolerance development, avoidance of negative withdrawal states and other factors that drivethem to therapy in their mid-adult years. This model suggests that interventions to reduce adolescent drinking will greatly improve abilities of many individuals andreduce overall lifetime alcoholism and addiction.
F. Crews et al. / Pharmacology, Biochemistry and Behavior 86 (2007) 189–199
neonatal anoxia: modulatory effects of an enriched environment. Psycho-
). Alcoholics have been found to see more fear in
pharmacology (Berl) 2006;184:155–65.
Andersen SL, Teicher MH. Delayed effects of early stress on hippocampal
facial expressions and animal studies have suggested these
alterations in fear response are the result of alcohol induced
Andersen SL, Thompson AT, Rutstein M, Hostetter JC, Teicher MH. Dopamine
deficits in associative learning Other
receptor pruning in prefrontal cortex during the periadolescent period in rats.
studies have found perseverative relearning deficits following a
rat model of binge drinking that relates to damage to
Andrucci GL, Archer RP, Pancoast DL, Gordon RA. The relationship of MMPI
and Sensation Seeking Scales to adolescent drug use. J Pers Assess
association cortex ). None of these
studies directly show a critical period of executive function
Arnett J. Reckless behavior in adolescence: a developmental perspective. Dev
development during adolescence that is disrupted by ethanol.
However, the findings of ethanol disruption of the critical
Bar-Peled O, Gross-Isseroff R, Ben-Hur H, Hoskins I, Groner Y, Biegon A. Fetal
period of visual cortical development, ethanol induced cortical
human brain exhibits a prenatal peak in the density of serotonin 5-HT1Areceptors. Neurosci Lett 1991;127:173–6.
neurotoxicity and ethanol induced alterations in executive
Baumrind D. A developmental perspective on adolescent rick taking in
function support the theory that disruption of frontal cortical
contemporary America. In: Irwin Jr CE, editor. Adolescent social behavior
development and executive function maturation occur in
and health. San Francisco, CA: Jossey-Bass; 1987. p. 93–125.
adolescent alcohol abusers (). It is possible that
Bender RA, Lauterborn JC, Gall CM, Cariaga W, Baram TZ. Enhanced CREB
adolescent alcohol abuse by disrupting impulse inhibition,
phosphorylation in immature dentate gyrus granule cells precedesneurotrophin expression and indicates a specific role of CREB in granule
attention and motivation promotes adult alcohol dependence
cell differentiation. Eur J Neurosci 2001;13:679–86.
and underlies the high risk of lifetime alcohol dependence
Berndt TJ. Developmental changes in conformity to peers and parents. Dev
found among those who begin drinking as adolescents. In total
the evidence does support a link between adolescent alcohol
Biglan A, Metzler CW, Wirt R, Ary D, Noell J, Ochs L, et al. Social and
abuse during a critical period of executive function develop-
behavioral factors associated with high-risk sexual behavior amongadolescents. J Behav Med 1990;13:245–61.
ment and an increased risk of lifetime alcohol dependence and
Blakemore SJ, Choudhury S. Development of the adolescent brain: implications
perhaps other psychopathology.
for executive function and social cognition. J Child Psychol Psychiatry2006;47:296–312.
Brooks-Kayal AR, Shumate MD, Jin H, Rikhter TY, Kelly ME, Coulter DA.
gamma-Aminobutyric acid(A) receptor subunit expression predicts func-tional changes in hippocampal dentate granule cells during postnatal
Adolescence represents an important period of brain
development. J Neurochem 2001;77:1266–78.
development, particularly for the cerebral cortex. There are
Burnet PW, Eastwood SL, Harrison PJ. Detection and quantitation of 5-HT1A
critical periods of development when the cortex is sensitive to
and 5-HT2A receptor mRNAs in human hippocampus using a reverse
environment induced changes in synaptic strength. Between the
transcriptase-polymerase chain reaction (RT-PCR) technique and their
ages of 10 and 25 years there are major changes in synaptic
correlation with binding site densities and age. Neurosci Lett 1994;178:85–9.
Cameron HA, Gould E. Adult neurogenesis is regulated by adrenal steroids in
receptors and density as well as myelination of frontal cortical
the dentate gyrus. Neuroscience 1994;61:203–9.
areas important for impulse control, goal setting, motivation,
Carlezon Jr WA, Duman RS, Nestler EJ. The many faces of CREB. Trends
interpersonal interactions, reasoning, assessment of rewards and
punishments in evaluating actions and other complex brain
Casey BJ, Trainor RJ, Orendi JL, Schubert AB, Nystrom LE, Cohen JD, et al. A
functions. Studies support the possibility that alterations in
pediatric functional MRI study of prefrontal activation during performanceof a Go-No-Go task. J Cogn Neurosci 1997;9:835–47.
adolescent synaptic remodeling create entrenchment of com-
CESAR. Despite declines in early initiation rates, many U.S. High School
mitted connections through altered pruning and focus of cortical
students still drink or smoke before age 13. Cesar Fax, vol. 15; 2006.
networks that underlie adult behavior repertoires. Heavy alcohol
Chambers RA, Taylor JR, Potenza MN. Developmental neurocircuitry of
consumption during adolescence disrupts cortical development
motivation in adolescence: a critical period of addiction vulnerability. Am J
altering higher executive functions in a manner that promotes
Choi S, Kellogg CK. Norepinephrine utilization in the hypothalamus of the male
continued impulsive behavior, alcohol abuse and risk of alcohol
rat during adolescent development. Dev Neurosci 1992;14:369–76.
Choi S, Weisberg SN, Kellogg CK. Control of endogenous norepinephrine
release in the hypothalamus of male rats changes over adolescent
development. Brain Res Dev Brain Res 1997;98:134–41.
Crews F, JG R, Chandler LJ. Glutamate and alcohol-induced neurotoxicity. In:
Herman BH, Frankenheim J, Litten R, Sheridan PH, Weight FF, Zukin SR,
This research is supported by NIAAA. We would also like to
editors. Glutamate and addiction. Humana Press; 2002. p. 357–73.
thank Melissa Mann for manuscript preparation.
Crews FT, Collins MA, Dlugos C, Littleton J, Wilkins L, Neafsey EJ, et al.
Alcohol-induced neurodegeneration: when, where and why? Alcohol ClinExp Res 2004;28:350–64.
Crews FT, Mdzinarishvili A, Kim D, He J, Nixon K. Neurogenesis in adolescent
brain is potently inhibited by ethanol. Neuroscience 2006;137:437–45.
Adriani W, Granstrem O, Macri S, Izykenova G, Dambinova S, Laviola G.
Csikszentmihalyi M, Larson R, Prescott S. The ecology of adolescent activity
Behavioral and neurochemical vulnerability during adolescence in mice:
and experience. J Youth Adolesc 1977;6:284–94.
studies with nicotine. Neuropsychopharmacology 2004;29:869–78.
Daval G, Verge D, Basbaum AI, Bourgoin S, Hamon M. Autoradiographic
Adriani W, Giannakopoulou D, Bokulic Z, Jernej B, Alleva E, Laviola G.
evidence of serotonin1 binding sites on primary afferent fibres in the dorsal
Response to novelty, social and self-control behaviors, in rats exposed to
horn of the rat spinal cord. Neurosci Lett 1987;83:71–6.
F. Crews et al. / Pharmacology, Biochemistry and Behavior 86 (2007) 189–199
De Bellis MD, Narasimhan A, Thatcher DL, Keshavan MS, Soloff P, Clark DB.
Gould E. Serotonin and hippocampal neurogenesis. Neuropsychopharmacology
Prefrontal cortex, thalamus, and cerebellar volumes in adolescents and
young adults with adolescent-onset alcohol use disorders and comorbid
Gould E, Woolf NJ, Butcher LL. Postnatal development of cholinergic neurons
mental disorders. Alcohol Clin Exp Res 2005;29:1590–600.
in the rat: I. Forebrain. Brain Res Bull 1991;27:767–89.
del Olmo E, Lopez-Gimenez JF, Vilaro MT, Mengod G, Palacios JM, Pazos A.
Grant BF, Dawson DA. Age of onset of drug use and its association with DSM-
Early localization of mRNA coding for 5-HT1A receptors in human brain
IV drug abuse and dependence: results from the National Longitudinal
during development. Brain Res Mol Brain Res 1998;60:123–6.
Alcohol Epidemiologic Survey. J Subst Abuse 1998;10:163–73.
Depue RA, Spoont MR. Conceptualizing a serotonin trait. A behavioral
Gregus A, Wintink AJ, Davis AC, Kalynchuk LE. Effect of repeated
dimension of constraint. Ann N Y Acad Sci 1986;487:47–62.
corticosterone injections and restraint stress on anxiety and depression-like
Dillon KA, Gross-Isseroff R, Israeli M, Biegon A. Autoradiographic analysis of
behavior in male rats. Behav Brain Res 2005;156:105–14.
serotonin 5-HT1A receptor binding in the human brain postmortem: effects
Guilarte T. The N-methyl-D-aspartate receptor: physiology and neurotoxicol-
of age and alcohol. Brain Res 1991;554:56–64.
ogy in the developing brain. In: Slikker Jr W, Chang LW, editors.
Dinopoulos A, Dori I, Parnavelas JG. The serotonin innervation of the basal
Handbook of developmental neurotoxicology. San Diego, CA: Academic
forebrain shows a transient phase during development. Brain Res Dev Brain
Press; 1998. p. 285–304.
Hachiya Y, Takashima S. Development of GABAergic neurons and their
Dori IE, Dinopoulos A, Parnavelas JG. The development of the synaptic
transporter in human temporal cortex. Pediatr Neurol 2001;25:390–6.
organization of the serotonergic system differs in brain areas with different
He J, Crews F. Neurogenesis Decreases during Brain Maturation from
functions. Exp Neurol 1998;154:113–25.
Adolescence to Adulthood. Pharmacol Biochem Behav 2007;86:327–33.
Draganski B, Gaser C, Busch V, Schuierer G, Bogdahn U, May A.
Hedner J, Lundell KH, Breese GR, Mueller RA, Hedner T. Developmental
Neuroplasticity: changes in grey matter induced by training. Nature
variations in CSF monoamine metabolites during childhood. Biol Neonate
2004;427: 311–2.
Draganski B, Gaser C, Kempermann G, Kuhn HG, Winkler J, Buchel C, et al.
Hensch TK. Critical period plasticity in local cortical circuits. Nat Rev Neurosci
Temporal and spatial dynamics of brain structure changes during extensive
learning. J Neurosci 2006;26:6314–7.
Huttenlocher PR. Synapse elimination and plasticity in developing human
Duka T, Gentry J, Malcolm R, Ripley TL, Borlikova G, Stephens DN, et al.
cerebral cortex. Am J Ment Defic 1984;88:488–96.
Consequences of multiple withdrawals from alcohol. Alcohol Clin Exp Res
Insel TR, Miller LP, Gelhard RE. The ontogeny of excitatory amino acid
receptors in rat forebrain-I. N-methyl-D-aspartate and quisqualate receptors.
Duman RS, Malberg J, Nakagawa S, D'Sa C. Neuronal plasticity and survival in
mood disorders. Biol Psychiatry 2000;48:732–9.
Jacobs BL, Praag H, Gage FH. Adult brain neurogenesis and psychiatry: a novel
Dyck RH, Cynader MS. Autoradiographic localization of serotonin receptor
theory of depression. Mol Psychiatry 2000;5:262–9.
subtypes in cat visual cortex: transient regional, laminar, and columnar
Johnson JS, Newport EL. Critical period effects in second language learning: the
distributions during postnatal development. J Neurosci 1993;13:4316–38.
influence of maturational state on the acquisition of English as a second
Ehardt C, Bernstein I. Patterns of affiliation among immature rhesus monkeys
language. Cognit Psychol 1989;21:60–99.
(Macaca mulatta). Am J Primatol 1987;13:255–69.
Johnston LD, O'Malley PM, Bachman JG, Schulenberg JE. Monitoring the
Fagen R. Exercise, play, and physical training in animals. Perspect Ethol
future, national survey results on drug use, 1975–2004. NIH Pub No 05-
5727 1 secondary school students; 2004.
Fassino M, Campbell B. The ontogeny of play in rats. Paper presented at
Kalsbeek A, Voorn P, Buijs RM, Pool CW, Uylings HB. Development of the
the meeting of the Eastern Psychological Association, New York, NY;
dopaminergic innervation in the prefrontal cortex of the rat. J Comp Neurol
Fehr C, Sander T, Tadic A, Lenzen KP, Anghelescu I, Klawe C, et al.
Keating D. Cognitive and brain development. Handbook of adolescent
Confirmation of association of the GABRA2 gene with alcohol dependence
psychology. 2nd ed.; 2004. p. 45–84.
by subtype-specific analysis. Psychiatr Genet 2006;16:9–17.
Kelley AE, Schochet T, Landry CF. Risk taking and novelty seeking in
Frantz K, Van Hartesveldt C. The locomotor effects of MK801 in the nucleus
adolescence: introduction to part I. Ann N Y Acad Sci 2004;1021:27–32.
accumbens of developing and adult rats. Eur J Pharmacol 1999a;368:
Kellogg CK. Early developmental modulation of GABAA receptor function.
Influence on adaptive responses. Perspect Dev Neurobiol 1998;5:219–34.
Frantz K, Van Hartesveldt C. Locomotion elicited by MK801 in developing and
Kellogg CK, Taylor MK, Rodriguez-Zafra M, Pleger GL. Altered stressor-
adult rats: temporal, environmental, and gender effects. Eur J Pharmacol
induced changes in GABAA receptor function in the cerebral cortex
of adult rats exposed in utero to diazepam. Pharmacol Biochem Behav
Gage FH. Mammalian neural stem cells. Science 2000;287:1433–8.
Galef JB. Mechanisms for the social transmission of food preferences from adult
Kellogg CK, Awatramani GB, Piekut DT. Adolescent development alters
to weaning rats. In: Barker LM, Best M, Domjan M, editors. Learning
stressor-induced Fos immunoreactivity in rat brain. Neuroscience 1998;
mechanisms in food selection. Baylor University Press; 1977. p. 123–48.
Geller B, Badner JA, Tillman R, Christian SL, Bolhofner K, Cook Jr EH.
Kempermann G. Why new neurons? Possible functions for adult hippocampal
Linkage disequilibrium of the brain-derived neurotrophic factor Val66Met
neurogenesis. J Neurosci 2002;22:635–8.
polymorphism in children with a prepubertal and early adolescent bipolar
Kostovic I. Structural and histochemical reorganization of the human prefrontal
disorder phenotype. Am J Psychiatry 2004;161:1698–700.
cortex during perinatal and postnatal life. Prog Brain Res 1990;85:223–39
Giedd JN. Structural magnetic resonance imaging of the adolescent brain. Ann
N Y Acad Sci 2004;1021:77–85.
Lauder JM. Ontogeny of the serotonergic system in the rat: serotonin as a
Giedd JN, Blumenthal J, Jeffries NO, Castellanos FX, Liu H, Zijdenbos A, et al.
developmental signal. Ann N Y Acad Sci 1990;600:297–313 [discussion
Brain development during childhood and adolescence: a longitudinal MRI
study. Nat Neurosci 1999;2:861–3.
Lauder JM, Bloom FE. Ontogeny of monoamine neurons in the locus coeruleus,
Gogtay N, Giedd JN, Lusk L, Hayashi KM, Greenstein D, Vaituzis AC, et al.
Raphe nuclei and substantia nigra of the rat. I. Cell differentiation. J Comp
Dynamic mapping of human cortical development during childhood through
early adulthood. Proc Natl Acad Sci U S A 2004;101:8174–9.
Le Moal M, Simon H. Mesocorticolimbic dopaminergic network: functional and
Golombek H, Kutcher S. Feeling states during adolescence. Psychiatr Clin
regulatory roles. Physiol Rev 1991;71:155–234.
North Am 1990;13:443–54.
Lewis DA, Cruz D, Eggan S, Erickson S. Postnatal development of prefrontal
Gordon JA, Stryker MP. Experience-dependent plasticity of binocular responses
inhibitory circuits and the pathophysiology of cognitive dysfunction in
in the primary visual cortex of the mouse. J Neurosci 1996;16:3274–86.
schizophrenia. Ann N Y Acad Sci 2004;1021:64–76.
F. Crews et al. / Pharmacology, Biochemistry and Behavior 86 (2007) 189–199
Malberg JE, Blendy JA. Antidepressant action: to the nucleus and beyond.
Sowell ER, Delis D, Stiles J, Jernigan TL. Improved memory functioning and
Trends Pharmacol Sci 2005;26:631–8.
frontal lobe maturation between childhood and adolescence: a structural
Malberg JE, Duman RS. Cell proliferation in adult hippocampus is decreased by
MRI study. J Int Neuropsychol Soc 2001;7:312–22.
inescapable stress: reversal by fluoxetine treatment. Neuropsychopharma-
Sowell ER, Thompson PM, Leonard CM, Welcome SE, Kan E, Toga AW.
Longitudinal mapping of cortical thickness and brain growth in normal
Malberg JE, Eisch AJ, Nestler EJ, Duman RS. Chronic antidepressant
children. J Neurosci 2004;24:8223–31.
treatment increases neurogenesis in adult rat hippocampus. J Neurosci
Spear LP. The adolescent brain and age-related behavioral manifestations.
Neurosci Biobehav Rev 2000;24:417–63.
Martin KC, Kandel ER. Cell adhesion molecules, CREB, and the formation of
Stanwood GD, McElligot S, Lu L, McGonigle P. Ontogeny of dopamine D3
new synaptic connections. Neuron 1996;17:567–70.
receptors in the nucleus accumbens of the rat. Neurosci Lett 1997;
Medina AE, Ramoa AS. Early alcohol exposure impairs ocular dominance
plasticity throughout the critical period. Brain Res Dev Brain Res
Steinberg L. Pubertal maturation and parent–adolescent distance: an evolution-
ary perspective. Advances in adolescent behavior and development. Sage
Medina AE, Krahe TE, Coppola DM, Ramoa AS. Neonatal alcohol exposure
Publications; 1989. p. 71–97.
induces long-lasting impairment of visual cortical plasticity in ferrets.
Strauss J, Barr CL, George CJ, King N, Shaikh S, Devlin B, et al. Association
J Neurosci 2003;23:10002–12.
study of brain-derived neurotrophic factor in adults with a history of
Medina AE, Krahe TE, Ramoa AS. Early alcohol exposure induces persistent
childhood onset mood disorder. Am J Med Genet B Neuropsychiatr Genet
alteration of cortical columnar organization and reduced orientation
selectivity in the visual cortex. J Neurophysiol 2005;93:1317–25.
Swann JW, Pierson MG, Smith KL, Lee CL. Developmental neuroplasticity:
Mehta AK, Ticku MK. Effect of chronic administration of ethanol on GABAA
roles in early life seizures and chronic epilepsy. Adv Neurol
receptor assemblies derived from alpha2-, alpha3-, beta2- and gamma2-
subunits in the rat cerebral cortex. Brain Res 2005;1031:134–7.
Tamm L, Menon V, Reiss AL. Maturation of brain function associated with
Merrick J, Kandel I, Birnbaum L, Hyam E, Press J, Morad M. Adolescent injury
response inhibition. J Am Acad Child Adolesc Psychiatry 2002;41:1231–8.
risk behavior. Int J Adolesc Med Health 2004;16:207–13.
Tanapat P, Hastings NB, Rydel TA, Galea LA, Gould E. Exposure to fox odor
Mohler H, Crestani F, Rudolph U. GABA(A)-receptor subtypes: a new
inhibits cell proliferation in the hippocampus of adult rats via an adrenal
pharmacology. Curr Opin Pharmacol 2001;1:22–5.
hormone-dependent mechanism. J Comp Neurol 2001;437:496–504.
Monti PM, Miranda Jr R, Nixon K, Sher KJ, Swartzwelder HS, Tapert SF, et al.
Tarazi FI, Baldessarini RJ. Comparative postnatal development of dopamine D
Adolescence: booze, brains, and behavior. Alcohol Clin Exp Res
(1), D(2) and D(4) receptors in rat forebrain. Int J Dev Neurosci 2000;18:
Morilak DA, Ciaranello RD. Ontogeny of 5-hydroxytryptamine2 receptor
Tarazi FI, Tomasini EC, Baldessarini RJ. Postnatal development of dopamine
immunoreactivity in the developing rat brain. Neuroscience 1993;55:869–80.
D4-like receptors in rat forebrain regions: comparison with D2-like
Munakata Y, Casey BJ, Diamond A. Developmental cognitive neuroscience:
receptors. Brain Res Dev Brain Res 1998a;110:227–33.
progress and potential. Trends Cogn Sci 2004;8:122–8.
Tarazi FI, Tomasini EC, Baldessarini RJ. Postnatal development of dopamine
Nixon K, Crews FT. Binge ethanol exposure decreases neurogenesis in adult rat
and serotonin transporters in rat caudate-putamen and nucleus accumbens
hippocampus. J Neurochem 2002;83:1087–93.
septi. Neurosci Lett 1998b;254:21–4.
Nurse S, Lacaille JC. Late maturation of GABA(B) synaptic transmission in area
Teicher MH. Limbic serotonin turnover plunges during puberty. Poster
CA1 of the rat hippocampus. Neuropharmacology 1999;38:1733–42.
presented at the meeting of the Society for Neuroscience, Miami Beach,
Obernier JA, White AM, Swartzwelder HS, Crews FT. Cognitive deficits and
CNS damage after a 4-day binge ethanol exposure in rats. Pharmacol
Teicher MH, Andersen SL, Hostetter Jr JC. Evidence for dopamine receptor
Biochem Behav 2002;72:521–32.
pruning between adolescence and adulthood in striatum but not nucleus
Prusky GT, Douglas RM. Developmental plasticity of mouse visual acuity. Eur J
accumbens. Brain Res Dev Brain Res 1995;89:167–72.
Thomas MJ, Beurrier C, Bonci A, Malenka RC. Long-term depression in the
Rosenberg DR, Lewis DA. Changes in the dopaminergic innervation of monkey
nucleus accumbens: a neural correlate of behavioral sensitization to cocaine.
prefrontal cortex during late postnatal development: a tyrosine hydroxylase
Nat Neurosci 2001;4:1217–23.
immunohistochemical study. Biol Psychiatry 1994;36:272–7.
Toga AW, Thompson PM, Sowell ER. Mapping brain maturation. Trends
Rosenzweig MR, Bennett EL. Psychobiology of plasticity: effects of training
and experience on brain and behavior. Behav Brain Res 1996;78:57–65.
Toscano CD, McGlothan JL, Guilarte TR. Lead exposure alters cyclic-AMP
Santarelli L, Saxe M, Gross C, Surget A, Battaglia F, Dulawa S, et al.
response element binding protein phosphorylation and binding activity in
Requirement of hippocampal neurogenesis for the behavioral effects of
the developing rat brain. Brain Res Dev Brain Res 2003;145:219–28.
antidepressants. Science 2003;301:805–9.
Toth G, Fekete M. 5-Hydroxyindole acetic excretion in newborns, infants and
Schramm NL, Egli RE, Winder DG. LTP in the mouse nucleus accumbens is
children. Acta Paediatr Hung 1986;27:221–6.
developmentally regulated. Synapse 2002;45:213–9.
Townshend JM, Duka T. Mixed emotions: alcoholics' impairments in the
Shaw P, Greenstein D, Lerch J, Clasen L, Lenroot R, Gogtay N, et al. Intellectual
recognition of specific emotional facial expressions. Neuropsychologia
ability and cortical development in children and adolescents. Nature
Ungless MA, Whistler JL, Malenka RC, Bonci A. Single cocaine exposure in
Shors TJ, Miesegaes G, Beylin A, Zhao M, Rydel T, Gould E. Neurogenesis in
vivo induces long-term potentiation in dopamine neurons. Nature
the adult is involved in the formation of trace memories. Nature
van Eden CG, Kros JM, Uylings HB. The development of the rat prefrontal
Silva AJ, Kogan JH, Frankland PW, Kida S. CREB and memory. Annu Rev
cortex. Its size and development of connections with thalamus, spinal cord
and other cortical areas. Prog Brain Res 1990;85:169–83.
Silveri MM, Spear LP. Decreased sensitivity to the hypnotic effects of ethanol
Vizuete ML, Venero JL, Traiffort E, Vargas C, Machado A, Cano J. Expression
early in ontogeny. Alcohol Clin Exp Res 1998;22:670–6.
of 5-HT7 receptor mRNA in rat brain during postnatal development.
Smith P. Does play matter? Functional and evolutionary aspects of animal and
Neurosci Lett 1997;227:53–6.
human play. Behav Brain Sci 1982;5:139–84.
Walker EF, Walder DJ, Reynolds F. Developmental changes in cortisol secretion
Sowell ER, Thompson PM, Holmes CJ, Batth R, Jernigan TL, Toga AW.
in normal and at-risk youth. Dev Psychopathol 2001;13:721–32.
Localizing age-related changes in brain structure between childhood and
Waters NS, Klintsova AY, Foster TC. Insensitivity of the hippocampus to
adolescence using statistical parametric mapping. Neuroimage 1999;9:
environmental stimulation during postnatal development. J Neurosci
F. Crews et al. / Pharmacology, Biochemistry and Behavior 86 (2007) 189–199
Wechsler H, Dowdall GW, Davenport A, Castillo S. Correlates of college
Zecevic N, Bourgeois JP, Rakic P. Changes in synaptic density in motor cortex
student binge drinking. Am J Public Health 1995;85:921–6.
of rhesus monkey during fetal and postnatal life. Brain Res Dev Brain Res
Weissenborn R, Duka T. Acute alcohol effects on cognitive function in social
drinkers: their relationship to drinking habits. Psychopharmacology (Berl)
Zehr JL, Todd BJ, Schulz KM, McCarthy MM, Sisk CL. Dendritic pruning of
the medial amygdala during pubertal development of the male Syrian
Wills TA, Vaccaro D, McNamara G. Novelty seeking, risk taking, and related
hamster. J Neurobiol 2006;66:578–90.
constructs as predictors of adolescent substance use: an application of
Zhu WJ, Roper SN. Brain-derived neurotrophic factor enhances fast excitatory
Cloninger's theory. J Subst Abuse 1994;6:1–20.
synaptic transmission in human epileptic dentate gyrus. Ann Neurol
Wilson M, Daly M. Competiveness, risk taking, and violence: the young male
syndrome. Ethol Sociobiol 1985:59–73.
Zou J, Crews F. CREB and NFkB transcription factors regulate sensitivity to
Yates T. Theories of cognitive development. In: Lewis M, editor. Child and
excitotoxic and oxidative stress induced neuronal cell death. Cell Mol
adolescent psychiatry. Baltimore: Williams and Wilkins; 1996. p. 134–55.
Yu ZY, Wang W, Fritschy JM, Witte OW, Redecker C. Changes in neocortical
and hippocampal GABAA receptor subunit distribution during brainmaturation and aging. Brain Res 2006;1099:73–81.
Source: https://tagasikooli.ee/wp-content/uploads/2015/03/adolescent-brain.pdf
le magazine Forum des Associations Enfance / Jeunesse • Quand le ciel de Royat • Une Ville active aux côtés • Une rentrée placée sous le signe s'embrase de mille feux ! des associations ! le magazine le magazine • Royat au cœur du réseau européen des villes thermales • Royat : 1ère station Thermale d'Auvergne • Royatonic : le sPA primé au niveau national
Invitation - Mission to Israel Friday 22 - Thursday 28 October 2010 from AUS $ 5,500 +GST ($5,500 + GST is the programming, administration and mission management fee. Approximate investment is $10,000 once accommodation, airfares and expenses are included) OPEN TO ALL INTERESTED PARTIES FROM AUSTRALIA AND NEW ZEALAND